 ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение
Как определить диапазон голоса - ваш вокал
Игровые автоматы с быстрым выводом
Как цель узнает о ваших желаниях прежде, чем вы начнете действовать. Как компании прогнозируют привычки и манипулируют ими
Целительная привычка
Как самому избавиться от обидчивости
Противоречивые взгляды на качества, присущие мужчинам
Тренинг уверенности в себе
Вкуснейший "Салат из свеклы с чесноком"
Натюрморт и его изобразительные возможности
Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д.
Как научиться брать на себя ответственность
Зачем нужны границы в отношениях с детьми?
Световозвращающие элементы на детской одежде
Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия
Как слышать голос Бога
Классификация ожирения по ИМТ (ВОЗ)
Глава 3. Завет мужчины с женщиной
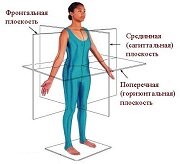
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
 Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу. Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу.
 Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар. Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.
| Понятие о веществах-агонистах и антагонистах
Вещества, которые при взаимодействии со специфическими рецепторами вызывают в них изменения, приводящие к биологическому эффекту, называют агонистами. Стимулирующее действие агониста на рецепторы может приводить к активации или угнетению функции клетки. Если агонист, взаимодействуя с рецепторами, вызывает максимальный эффект, то это полный агонист. В отличие от последнего частичные агонисты при взаимодействии с теми же рецепторами не вызывают максимального эффекта.
Вещества, связывающиеся с рецепторами, но не вызывающие их стимуляции, называют антагонистами. Их внутренняя активность равна нулю. Их фармакологические эффекты обусловлены антагонизмом с эндогенными лигандами (медиаторами, гормонами), а также с экзогенными веществами-агонистами. Если они оккупируют те же рецепторы, с которыми взаимодействуют агонисты, то речь идет о конкурентных антагонистах; если другие участки макромолекулы, не относящиеся к специфическому рецептору, но взаимосвязанные с ним, то говорят о неконкурентных антагонистах.
Если вещество действует как агонист на один подтип рецепторов и как антагонист - на другой, оно обозначается как агонист-антагонист.
Выделяют и так называемые неспецифические рецепторы, связываясь с которыми вещества не вызывают возникновения эффекта (белки плазмы крови, мукополисахариды соединительной ткани); их еще называют местами неспецифического связывания веществ.
Взаимодействие "вещество-рецептор" осуществляется за счет межмолекулярных связей. Один из наиболее прочных видов связи - ковалентная связь. Она известна для небольшого количества препаратов (некоторые противобластомные вещества). Менее стойкой является более распространенная ионная связь, типичная для ганглиоблокаторов и ацетилхолина. Важную роль играют вандерваальсовы силы (основа гидрофобных взаимодействий) и водородные связи.
В зависимости от прочности связи "вещество-рецептор" различают обратимое действие, характерное для большинства веществ, и необратимое действие (в случае ковалентной связи).
Если вещество взаимодействует только с функционально однозначными рецепторами определенной локализации и не влияет на другие рецепторы, то действие такого вещества считают избирательным. Основой избирательности действия является сродство (аффинитет) вещества к рецептору.
Другой важной мишенью лекарственных веществ являются ионные каналы. Особый интерес представляет поиск блокаторов и активаторов Са2+-каналов с преимущественным влиянием на сердце и сосуды. В последние годы большое внимание привлекают вещества, регулирующие функцию К+-каналов.
Важной мишенью многих лекарственных веществ являются ферменты. Например, механизм действия нестероидных противовоспалительных средств обусловлен ингибированием циклооксигеназы и снижением биосинтеза простогландинов. Антибластомный препарат метотрексат блокирует дигидрофолатредуктазу, препятствуя образованию тетрагидрофолата, необходимого для синтеза пуринового нуклеотида-тимидилата. Ацикловир ингибирует вирусную ДНК-полимеразу.
Еще одна возможная мишень лекарственных средств - транспортные системы для полярных молекул, ионов и мелких гидрофильных молекул. Одно из последних достижений в этом направлении - создание ингибиторов пропионового насоса в слизистой оболочке желудка (омепразол).
Важной мишенью многих лекарственных веществ считаются гены. Исследования в области генной фармакологии получают все более широкое распространение.
12.Симпатический отдел вегетативной нервной системы и его роль в регуляции жизнедеятельности организма. Морфологически и функционально выделяют два отдела вегетативной нервной системы : симпатическую и парасимпатическую нервные системы.
Симпатическая система мобилизует силы организма в экстренных ситуациях, увеличивает трату энергетических ресурсов; парасимпатическая - способствует восстановлению и накоплению энергетических ресурсов.
Активность симпатической нервной системы и секреция адреналина мозговым веществом надпочечников связаны друг с другом, но не всегда изменяются в одинаковой степени. Так, при особо сильной стимуляции симпатоадреналовой системы (например, при общем охлаждении или интенсивной физической нагрузке ) возрастает секреция адреналина, усиливая действие симпатической нервной системы. В других же ситуациях симпатическая активность и секреция адреналина могут быть независимы. В частности, в ортостатической реакции участвует в основном симпатическая нервная система, а в реакции на гипогликемию - мозговое вещество надпочечников. Окончания симпатических нервов образуют в иннервируемых тканях сплетения. Весь норадреналин , содержащийся в тканях, находится в симпатических окончаниях. В органах с обильной симпатической иннервацией его концентрация достигает 1-2мкг/г. Норадреналин в симпатических окончаниях, как и в мозговом веществе надпочечников , содержится в пузырьках. МАО , локализованная в митохондриях симпатических окончаний, играет важную роль в регуляции местной концентрации норадреналина ( рис. 70.2 ).
Катехоламины , содержащиеся в пузырьках, защищены от действия МАО, но свободные катехоламины в цитоплазме дезаминируются с образованием неактивных метаболитов.
Высвобождение норадреналина из симпатических окончаний запускается поступающим в эти окончания потенциалом действия.
Тела преганглионарных симпатических нейронов сосредоточены в промежуточном и боковом сером веществе ( интермедиолатеральном столбе ) грудных и поясничных сегментов спинного мозга ( рис. 41.1 и рис. 41.2 ). Некоторые нейроны обнаружены в сегментах С8. Наряду с локализацией в интермедиолатеральном столбе выявлена локализация преганглионарных симпатических нейронов также в боковом канатике, промежуточной области спинного мозга и пластине X ( дорсальнее центрального канала).
Большинство симпатических ганглиев удалено от иннервируемых органов, и поэтому от них идут длинные постганглионарные аксоны .
У большинства преганглионарных симпатических нейронов тонкие миелинизированные аксоны - В-волокна . Однако некоторые аксоны относятся к немиелинизированным С-волокнам. Скорость проведения по этим аксонам колеблется от 1 до 20 м/с. Они покидают спинной мозг в составе передних корешков и белых соединительных ветвей и оканчиваются в парных паравертебральных ганглиях или непарных превертебральных ганглиях . Посредством нервных веточек паравентебральные ганглии соединены в симпатические стволы , идущие по обе стороны позвоночника от основания черепа до крестца. От симпатических стволов отходят более тонкие немиелинизированные постганглионарные аксоны , которые либо направляются к периферическим органам в составе серых соединительных ветвей, либо образуют специальные нервы, идущие к органам головы, грудной, брюшной и тазовой полостей. Постганглионарные волокна от превертебральных ганглиев (чревного, верхнего и нижнего брызжеечных) идут через сплетения или в составе особых нервов к органам брюшной полости и органам полости таза.
Преганглионарные аксоны покидают спинной мозг в составе переднего корешка и через белые соединительные ветви входят в паравертебральный ганглий на уровне того же сегмента. Белые соединительные ветви есть только на уровнях Th1-L2. Преганглионарные аксоны оканчиваются синапсами в этом ганглии или, пройдя через него, входят в симпатический ствол ( симпатическую цепочку ) паравертебральных ганглиев либо во внутренностный нерв ( рис. 41.2 ).
В составе симпатической цепочки преганглионарные аксоны направляются рострально либо каудально к ближайшему или удаленному паравертебральному ганглию и там образуют синапсы . Выйдя из него, аксоны идут к спинальному нерву обычно через серую соединительную ветвь, которая есть у каждого из 31 пары спинальных нервов. В составе периферических нервов постганглионарные аксоны поступают к эффекторам кожи (пилоэректорным мышцам, кровеносным сосудам , потовым железам ), мышц, суставов. Как правило, постганглионарные аксоны немиелинизированы ( С-волокна ), хотя есть исключения. Различия между белыми и серыми соединительными ветвями зависят от относительного содержания в них миелинизированных и немиелинизированных аксонов.
В составе внутренностного нерва преганглионарные аксоны часто идут к превертебральному ганглию , где образуют синапсы, либо они могут проходить через ганглий, оканчиваясь в более удаленном ганглии. Некоторые из них, идущие в составе внутренностного нерва, оканчиваются непосредственно на клетках мозгового вещества надпочечников.
Симпатическая цепочка тянется от шейного до копчикового уровня спинного мозга. Она выполняет роль распределительной системы, позволяя преганглионарным нейронам, которые располагаются только в грудных и верхних поясничных сегментах, активировать постганглионарные нейроны, снабжающие все сегменты тела. Однако паравертебральных ганглиев меньше, чем спинальных сегментов, так как некоторые ганглии сливаются в процессе онтогенеза . Например, верхний шейный симпатический ганглий состоит из слившихся ганглиев С1-С4, средний шейный симпатический ганглий - из С5-С6, а нижний шейный симпатический ганглий - из С7-С8. Звездчатый ганглий образован слиянием нижнего шейного симпатического ганглия с ганглием Th1. Верхний шейный ганглий обеспечивает постганглионарную иннервацию головы и шеи, а средний шейный и звездчатый - сердца, легких и бронхов.
Обычно аксоны преганглионарных симпатических нейронов распределяются к ипсилатеральным ганглиям и, следовательно, регулируют вегетативные функции на той же стороне тела. Важное исключение - двусторонняя симпатическая иннервация кишечника и органов таза. Так же как двигательные нервы скелетных мышц, аксоны преганглионарных симпатических нейронов, относящиеся к определенным органам, иннервируют несколько сегментов. Так, преганглионарные симпатические нейроны, обеспечивающие симпатические функции областей головы и шеи, находятся в сегментах С8-Th5, а те, которые относятся к надпочечникам, - в Th4-Th12.
К эффекторам, снабжаемым симпатической системой , относятся гладкие мышцы всех органов ( сосудов , органов брюшной полости , выделительных органов , легких , зрачка ), сердце и некоторые железы ( потовые , слюнные и пищеварительные железы ). Кроме того, симпатические постганглионарные волокна иннервируют клетки подкожной жировой клетчатки , печени , и, возможно, канальцы почек . 13.Парасимпатический отдел вегетативной нервной системы и его роль в регуляции жизнедеятельности организма. Центрами парасимпатического отдела автономной нервной системы являются ядра, находящиеся в среднем мозге (III пара черепно-мозговых нервов), продолговатом мозге (VII, IX и X пары черепно-мозговых нервов) и крестцовом отделе спинного мозга (ядра тазовых внутренних нервов). От среднего мозга отходят преганглионарные волокна парасимпатических нервов, которые входят в состав глазодвигательного нерва.
Из продолговатого мозга выходят преганглионарные волокна, идущие в составе лицевого, языкоглоточного и блуждающих нервов. От крестцового отдела спинного мозга отходят преганглионарные парасимпатические волокна, которые входят в состав тазового нерва. Ганглии парасимпатической нервной системы располагаются вблизи иннерви-руемых органов или внутри них. Поэтому преганглионарные волокна парасимпатического отдела длинные, а постганглионарные волокна короткие по сравнению с волокнами симпатического отдела.
В окончаниях как преганглионарных, так и большинства постганглионарных волокон вырабатывается ацетилхолин. Парасимпатические волокна иннервируют, как правило, только определенные части тела, которые имеют также симпатическую, а иногда и внутриорганную иннервацию. Парасимпатическая нервная система не иннервирует скелетные мышцы, головной мозг, гладкие мышцы кровеносных сосудов, за исключением сосудов языка, слюнных желез, половых желез и коронарных артерий, органы чувств и мозговое вещество надпочечников. Постган-глионарпые парасимпатические волокна иннервируют глазные мышцы, слезные и слюнные железы, мускулатуру и железы пищеварительного тракта, трахею, гортань, легкие, предсердия, выделительные и половые органы.
При возбуждении парасимпатических нервов тормозится работа сердца (отрицательные хроно-, ино-, дромо- и батмотроп-ное действия), повышается тонус гладкой мускулатуры бронхов, в результате чего уменьшается их просвет, сужается зрачок, стимулируются процессы пищеварения (моторика и секреция), обеспечивая тем самым восстановление уровня питательных веществ в организме, происходит опорожнение желчного пузыря, мочевого пузыря, прямой кишки. Действие парасимпатической нервной системы направлено на восстановление и поддержание постоянства состава внутренней среды организма, нарушенного в результате возбуждения симпатической нервной системы. Парасимпатическая нервная система выполняет в организме трофотроп-ную функцию 14.Механизм распространения возбуждения по миелинизированным и немиелинизированным нервным волокнам. Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки — нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 2.3). Чехол прерывается примерно через каждые 0,5-1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис. 2.9 показано, как это происходит. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии. 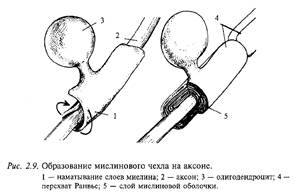
Образование миелинового чехла на аксоне
Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга..
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре. 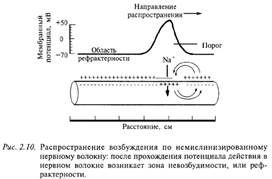
Распространение возбуждения по немиелинизированному нервному волокну: после прохождения потенциала действия в нервном волокне возникает зона невозбудимости, или рефрактерности
Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис. 2.10 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы,-выходящий ток, который деполяризует соседний невозбужденный участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность — зона невозбудимости. Нам уже известно, что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100 — 120 м/с, должны иметь диаметр около 1000 мкм (1 мм).
У млекопитающих животных природа сохранила немиелинизированными только те возбуждение о боли, температуре, управляют медленно работающими внутренними органами мочевым волокна, которые проводят органами — мочевым пузырем, кишечником и пр. Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рис. 2.11 показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые находятся на стыке между двумя участками миелина
Распространение возбуждения по немиелинизированному нервному волокну: потенциалы действия возникают только в перехватах Ранвье. 15.Классификация нервных волокон. Факторы, определяющие скорость проведения возбуждения по аксонам.
Нервные волокна классифицируются по:
1.длительности потенциала действия;
2.строению (диаметру) волокна;
3.скорости проведения возбуждения.
Выделяют следующие группы нервных волокон:
1.группа А (альфа, бета, гамма, дельта) - самый короткий потенциал действия, самая толстая миелиновая оболочка, самая высокая скорость проведения возбуждения;
2.группа В - миелиновая оболочка менее выражена;
3.группа С - без миелиновой оболочки.
Классификация Эрлангера-Гассера Является наиболее полной классификацией нервных волокон по скорости проведения нервного импульса. | Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация | | Aα | Афферентные — мышечные веретёна, сухожильные органы; эфферентные — скелетные мышцы | 10-20 | 60-120 | + | | Aβ | Афферентные — тактильное чувство; коллатерали Aα волокон к интрафузальным мышечным волокнам | 7-15 | 40-90 | + | | Aγ | Эфферентные — мышечные веретёна | 4-8 | 15-30 | + | | Aδ | Афферентные — температура, быстрое проведение боли | 3-5 | 5-25 | + | | B | Симпатические, преганглионарные; постганглионарные волокна цилиарного ганглия | 1-3 | 3-15 | прерывистая | | C | Симпатические, постганглионарные; афферентные — медленное проведение боли | 0,3-1 | 0,5-2 | - | 16.Классификация медиаторов и модуляторов ЦНС. По химической структуре медиаторы подразделяют на:
• моноамины (адреналин, норадреналин, ацетилхолин и др.);
• аминокислоты (гамма-аминомасляная кислота (ГАМК), глутамат, глицин, таурин);
• пептиды (эндорфин, нейротензин, бомбезин, энкефалин и др.);
• прочие медиаторы (NO , АТФ).
Амбивалентность действия медиаторов проявляется в том, что один и тот же медиатор в разных синапсах может оказывать различное действие на эффекторную клетку. Результат действия медиатора на постсинаптическую мембрану зависит от того, какие рецепторы и ионные каналы в ней находятся. Если медиатор открывает в постсинаптической мембране Na+ -каналы, то это приводит к развитию ВПСП, если K+ - или Cl – -каналы, то развивается ТПСП. Вследствие этого термины «возбуждающий медиатор» и «тормозный медиатор» неправомерны; следует говорить лишь о возбуждающих и тормозных синапсах.
В синаптическом окончании наряду с медиатором могут синтезироваться и высвобождаться одно или несколько химических веществ. Эти соединения, действуя на постсинаптичекую мембрану, могут повышать или снижать ее возбудимость. Поскольку сами по себе они не могут вызвать возбуждение постсинаптической мембраны, их называют модуляторами синаптической передачи (нейромодуляторами). Большинство нейромодуляторов представляют собой пептиды. 17.Понятие о медиаторах и модуляторах. Критерии (признаки) медиатора. Не сразу бывает понятно, чем же именно отличаются друг от друга нейромедиаторыи нейромодуляторы. Оба вида этих управляющих веществ содержатся в синаптических пузырьках пресинаптических окончаний и выбрасываются в синаптическую щель. Они относятся к нейротрансмиттерам - передатчикам управляющих сигналов. Медиаторы и модуляторы отличаются друг от друга по нескольким признакам. Это поясняет размещённый здесь оригинальный рисунок. Попробуйте найти на нём эти отличия...
Говоря об общем числе известных медиаторов, можно перечислить от десятка до сотни химических веществ. Критерии нейромедиаторов
1. Вещество выделяется из нейрона при его активации.
2. В клетке присутствуют ферменты для синтеза данного вещества.
3. В соседних клетках (клетках-мишенях) выявляются белки-рецепторы, активируемые данным медиатором.
4. Фармакологический (экзогенный) аналог имитирует действие медиатора.
Иногда медиаторы объединяют с модуляторами, то есть веществами, которые прямо не участвуют в процессе передачи сигнала (возбуждения или торможения) от нейрона к нейрону, но могут, однако, этот процесс существенно усиливать или ослаблять. Первичные медиаторы - это те, которые действуют непосредственно на рецепторы постсинаптической мембраны.
Сопутствующие медиаторы и медиаторы-модуляторы - могут запускать каскад ферментативных реакций, которые, например, изменяют чувствительность рецептора к первичному медиатору.
Аллостерические медиаторы - могут участвовать в кооперативных процессах взаимодействия с рецепторами первичного медиатора.
Важнейшим отличием медиаторов от модуляторов считается то, что медиаторы способны передавать возбуждение или наводить торможение на клетку-мишень, в то время как модуляторы лишь подают сигнал к началу метаболических процессов внутри клетки.
Медиаторы связываются с ионотропными молекулярными рецепторами, которые являются наружной частью ионных каналов. Поэтому медиаторы могут открывать ионные каналы и тем самым запускать трансмембранные потоки ионов. Соответственно, входящие в ионные каналы положительные ионы натрия или кальция вызывают деполяризацию (возбуждение), а входящие отрицательные ионы хлора - гиперполяризацию (торможение). Ионотропные рецепторы вместе со своими каналами сосредоточены на постсинаптической мембране. Всего известно примерно 20 видов медиаторов.
Модуляторы же связываются с метаботропными молекулярными рецепторами, которые сидят отдельно от ионных каналов на любом месте мембраны. С внутренней стороны мембраны к этим рецепторам присоединяются G-белки. Когда модулятор связывается с метаботропным рецептором, то G-белок активируется и запускает каскад биохимических реакций внутри клетки. Таким способом модулируется (т.е. изменяется) внутреннее состояние клетки. Вот почему эти вещества и называются модуляторами. В отличие от медиаторов, известно намного больше видов модуляторов - более 600 по сравнению с 20 медиаторами. Практически все модуляторы являются по химическому строению нейропептидами, т.е. аминокислотными цепочками, более короткими, чем белки. Интересно, что некоторые медиаторы "по совместительству" могут играть и роль модуляторов, т.к. к ним имеются метаботропные рецепторы. Таковы, например, серотонин и ацетилхолин.
Механизм внутриклеточных эффектов модуляторов, осуществляющих медленную синаптическую передачу, был раскрыт в исследованиях Пола Грингарда (Paul Greengard). Он продемонстрировал, что, помимо классических эффектов, реализующихся через ионотропные рецепторы и непосредственное изменение электрических мембранных потенциалов, многие нейротрансмиттеры (катехоламины, серотонин и многие нейропептиды) оказывают влияние на биохимические процессы в цитоплазме нейронов. Именно этими метаботропными эффектами и обусловлено необычно медленное действие таких трансмиттеров и их длительное модулирующее влияние на функции нервных клеток. Поэтому именно нейромодуляторы вовлечены в обеспечение сложных состояний нервной системы - эмоций, настроений, мотиваций, а не в передачу быстрых сигналов для восприятия, движения, речи и т.д. 18.Дофаминэргическая система мозга.
В этой нейрохимической системе мозга различают 7 отдельных подсистем: нигростриатную, мезокортикальную, мезолимбическую, тубероинфундибулярную, инцертогипоталамическую, диенцефалоспинальную и ретинальную. Из них первые 3 являются основными.
Тела нейронов нигростриатной, мезокортикальной и мезолимбической систем расположены на уровне среднего мозга и образуют комплекс нейронов чёрной субстанции (substantia nigra) и вентрального поля покрышки. Они составляют непрерывную клеточную сеть, проекции которой частично перекрываются, поскольку аксоны этих нейронов идут вначале в составе одного крупного тракта (медиального пучка переднего мозга), а оттуда расходятся в разные мозговые структуры. Формирование нигростриатной, мезолимбической и мезокортикальной систем определяется областями, где оканчиваются аксоны дофаминергических нейронов, т.е. локализацией их проекций. Некоторые авторы объединяют мезокортикальную и мезолимбическую подсистемы в единую систему. Более обоснованным является выделение мезокортикальной и мезолимбической подсистем соответственно проекциям в лобную кору и лимбические структуры мозга.
Нигростриатная подсистема
Нигростриатный тракт является самым мощным в дофаминергической системе мозга. Аксонами нейронов этого тракта выделяется около 80 % мозгового дофамина. Тела дофаминовых нейронов, образующих этот путь, находятся в основном в компактной части черной субстанции, но часть волокон берет начало также от нейронов латерального отдела вентрального поля покрышки среднего мозга.
Мезокортикальная подсистема
Тела нейронов, образующих мезокортикальный тракт, находятся в вентральной части покрышки среднего мозга, а основные проекции этих нейронов достигают лобной (преимущественно префронтальной, поле 10 по Бродману - рис. 9) коры. Соответствующие окончания расположены в основном в глубоких слоях лобной коры (V-VI). Мезокортикальная дофаминовая система оказывает большое влияние на активность нейронов, образующих корково-корковые, корково-таламичес-кие и корково-стриатные пути.
Мезолимбическая подсистема
Источники дофаминергических проекций, т.е. тела нейронов этой системы, расположены в вентральном поле покрышки среднего мозга и частично в компактной части черной субстанции. Их отростки идут в поясную извилину, энториальную кору, миндалину, обонятельный бугорок, аккумбентное ядро, гиппокамп, парагиппокампальную извилину, перегородку и другие структуры лимбической системы мозга. Имея обширные связи, Мезолимбическая система опосредовано проецируется также на лобную кору и гипоталамус. Это определяет широкие функции мезолимбической системы, которая участвует в механизмах памяти, эмоций, обучения и нейроэндокринной регуляции.
Другие тракты
Тубероинфундибулярный тракт образован аксонами нейронов, расположенных в аркуатном ядре гипоталамуса. Отростки таких нейронов достигают наружного слоя срединного возвышения. Этот тракт осуществляет контроль секреции пролактина. Дофамин тормозит его секрецию и поэтому содержание пролактина в плазме крови служит косвенным показателем функции дофаминергической системы мозга, что часто используют для оценки влияния на нее психофармакологических средств. Инцертогипоталамический тракт начинается от zona incerta и оканчивается в дорсальном и переднем отделах медиального таламуса, а также в перивентрикулярной области. Он принимает участие в нейроэндокринной регуляции. Источником проекций диенцефалоспинального тракта являются нейроны заднего гипоталамуса, отростки которых достигают задних рогов спинного мозга. Ретинальный тракт расположен в пределах сетчатки глаза. Особенности этого тракта делают его среди других дофаминергических трактов достаточно автономным. 19.Ацетилхолин, его рецепторы и роль как медиатора в периферической, вегетативной и центральной нервной системе. Периферическое никотиноподобное дейсвие ацетилхолина связано с его участием в передаче нервных импульсов с преганглионарных волокон на постганглионарные в вегетативных узлах, а также с двигательных нервов на поперечнополосатую мускулатуру. В малых дозах он является физиологическим передатчиком нервного возбуждения, в больших дозах может вызвать стойкую деполяризацию в области синапсов и блокировать передачу возбуждения. Ацетилхолину принадлежит также важная роль как медиатору ЦНС. Он участвует в передаче импульсов в разных отделах мозга, при этом малые концентрации облегчают, а большие - тормозят синаптическую передачу. Изменения в обмене ацетилхолина могут привести к нарушению функций мозга. Некоторые центральнодействующие антагонисты ацетилхолина (см. Амизил) являются психотропными препаратами (см. также Атропин). Передозировка антагонистов ацетилхолина может вызвать нарушения высшей нервной деятельности (оказывать галлюциногенный эффект и др.). Для применения в медицинской практике и для экспериментальных исследований выпускается ацетилхолин-хлорид (Acetylcholini chloridum). Синонимы: Acetylchlolinum chloratum, Acecoline, citocholine, Miochol и др. Бесцветные кристаллы или белая кристаллическая масса. Расплывается на воздухе. Легко растворим в воде и спирте. 20. Норадренэргическая система мозга. Сходство и различие между адреналином и норадреналином. 1.Норадренергическая система. Источником норадренергических путей в мозге являются группы клеток, расположенных в мозговом стволе и ретикулярной формации. Они включают клетки голубого пятна (locus ceruleus), вентромедиальной части покрышки и др. Отростки таких клеток сильно разветвлены и коллатеризированы. Области, на которые распространяются восходящие проекции этих клеток, захватывают ствол мозга, гипоталамус, таламус и разные отделы коры, а нисходящие достигают спинного мозга. Восходящие норадренергические проекции являются компонентом восходящих активирующих систем.
Адренергические рецепторы разделяются на а- и J3-, а последние на (3,-и (3> Рецепторы В, локализуются на нейроне, а (39 - на клетках глии и сосудовАгонистом |3|-рецепторов является норадреналин, а В2-рецепторы более чувствительны к адреналину.
Рецепторы типов at и а2 хорошо изучены фармакологически. Специфические ингибиторы агрецепторов обладают антигипертензивными свойствами, а2-рецепторы в большой степени определяют активность центральной и периферической адренергической систем. Пресинаптические ^-рецепторы на норадренергических терминалях тормозят выделение норад-реналина, имея отношение и к регуляции кровяного давления. Об этом свидетельствует, в частности, влияние клонидина, который будучи антиги-пертензивным средством уменьшает также симптомы абстиненции при алкоголизме и наркоманиях. 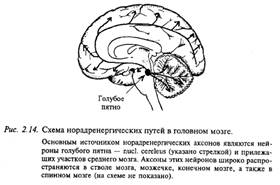
Схема норадренергических путей в головном мозге
Основным источником норадренергических аксонов являются нейроны голубого пятна и прилежащих участков среднего мозга (рис. 2.14). Аксоны этих нейронов широко распространяются в мозговом стволе, мозжечке, в больших полушариях. В продолговатом мозге крупное скопление норадренергических нейронов находится в вентролатеральном ядре ретикулярной формации. В промежуточном мозге (гипоталамусе) норадренергические нейроны наряду с дофаминергическими нейронами входят в состав гипоталамо-гипофизарной системы. Норадренергические нейроны в большом количестве содержатся в нервной периферической системе. Их тела лежат в симпатической цепочке и в некоторых интрамуральных ганглиях.
2.Адреналин вызывает те же эффекты, как и норадреналин, но есть некоторые различия. Во-первых, адреналин в связи с более выраженной стимуляцией бета-рецепторов сильнее влияет на сердце, чем норадреналин. Во-вторых, адреналин вызывает только слабое сужение кровеносных сосудов в мышцах по сравнению с гораздо более сильным сужением, вызываемым норадреналином. Поскольку мышечные сосуды составляют большую часть сосудов тела, это отличие особенно важно, т.к. норадреналин значительно увеличивает общее периферическое сопротивление и повышает артериальное давление, тогда как адреналин поднимает давление в меньшей степени, но больше увеличивает сердечный выброс.
Третье различие между действием адреналина и норадреналина связано с их воздействием на тканевой метаболизм. Адреналин имеет в 5-10 раз более длительный метаболический эффект, чем норадреналин. Действительно, адреналин, секретируемый мозговым веществом надпочечников, может увеличивать интенсивность метаболизма всего тела более чем на 100% выше нормы, таким образом увеличивая активность и возбудимость организма. Он также увеличивает скорость других метаболических явлений, например гликогенолиза в печени и в мышцах и освобождения глюкозы в кровь.
Итак, стимуляция мозгового вещества вызывает освобождение гормонов адреналина и но-радреналина, которые совместно имеют те же эффекты в организме, что и прямая симпатическая стимуляция; кроме того, эффекты гормонов значительно продолжительнее и длятся 2-4 мин после окончания стимуляции. 21.Нейропептиды как медиаторы и модуляторы в ЦНС: основные представители и их функции. Нейропептиды -биологически активные соединения, синтезируемые главным образом в нервных клетках. Участвуют в регуляции обмена веществ и поддержании гомеостаза, воздействуют на иммунные процессы, играют важную роль в механизмах памяти, обучения, сна и др. Могут действовать как медиаторы и гормоны. Часто один и тот же нейропептид способен выполнять различные функции (напр., ангиотензин, энкефалины, эндорфины). Используются в медицине как лекарственные средства.
За последние годы, после того как в мозге был обнаружен новый класс химических соединений – нейропептиды, число известных систем химических посредников в головном мозге резко возросло. Нейропептиды - биологически активные соединения, синтезируемые главным образом в нервных клетках. Участвуют в регуляции обмена веществ и поддержании гомеостаза, воздействуют на иммунные процессы, играют важную роль в механизмах памяти, обучения, сна и др. Могут действовать как медиаторы и гормоны. Часто один и тот же нейропептид способен выполнять различные функции (напр., ангиотензин, энкефалины, эндорфины). Используются в медицине как лекарственные средства.
Нейропептиды присутствуют в немиелинизированных волокнах С-типа и небольших миелинизированных А-дельта-типа волокнах и синтезируются клетками дорзальных рогов ганглиев, а затем транспортируются вдоль аксонов в нервные окончания, где они накапливаются в плотных везикулах. Вначале было изучено влияние нейропептидов на сосудистый тонус. Однако впоследствии было обнаружено, что некоторые из них генерируют и поддерживают воспалительный процесс, называемый «нейрогенным». Выделяют следующие семейства нейропептидов: 22.Система опиоидных пептидов в нервной системе: рецепторы, механизм действия, связь с наркотической зависимостью. Опиоидная система регуляции боли включает опиатные рецепторы и опиоидные пептиды.
Опиатные рецепторы представлены миорецепторами (д-рецепторами), сигмарецепторами (о-рецепторами) и каппа-рецепторами (к-рецепторами). Эти рецепторы присутствуют во всех структурах НЦС, главным образом — в основных релейных станциях афферента-ции ноцицептивной импульсации (желатинозной субстанции задних рогов спинного мозга, гигантоклеточном ядре продолговатого мозга, центральном сером околоводопроводном веществе и покрышке среднего мозга, синем пятне, чёрной субстанции, красном ядре, ядрах ретикулярной формации, гипоталамуса, таламуса, лимбических структур, а также в корковых центрах боли). В одних образованиях ЦНС (фронтальной коре, лимбических структурах) опиатных рецепторов во много раз больше, чем в других (теменной, височной и затылочной долях).
Опиоидные пептиды представлены эндорфинами и энкефалинами.
- Эндорфины (а, р, у) выполняют роль медиаторов АНЦС и имеют в ЦНС более узкую локализацию их синтеза и действия. В гипоталамусе, гипофизе, перегородке мозга, среднем мозге, таламусе эндорфинов значительно больше, чем энкефалинов.
- Энкефалины играют роль и медиаторов, и модуляторов. Они имеют более обширную локализацию их синтеза и действия в ЦНС. Причём энкефалины оказывают общее угнетающее действие на активность различных нейронов ЦНС, снижая их реакции на любой сенсорный раздражитель. В отличие от эндорфинов, энкефалины оказывают более слабое угнетающее действие на ЦНС.
Эндорфины, как и экзогенный морфин, оказывают угнетающее влияние на ноцицептивные синаптические входы, а также активируют большинство нейронов АНЦС. Таким образом, эндорфины снижают болевую чувствительность и болевое ощущение не только за счёт угнетения структур НЦС, но и за счёт активизации структур АНЦС. Следует подчеркнуть, что эндогенная опиоидная система с участием эндорфинов, энкефалинов и опиатных рецепторов — надёжный регулятор и контролёр интенсивности ноцицептивного возбуждения. С увеличением интенсивности ноцицептивной импульсации опиоидная система активизируется в большей степени. Ненарушенная опиоидная система всегда находится в активном состоянии и способна ограничивать степень возбуждения различных сенсорных, в том числе и болевых структур.
Отмечено, что содержание опиоидных пептидов в биологических средах организма, особенно в структурах АНЦС, как и активность опиатных рецепторов в различных образованиях этой системы подвержены суточным колебаниям. Этим, вероятно, и можно объяснить суточные ритмы болевой чувствительности.
Показано также, что опиатные рецепторы образуют обратимую связь с наркотическими анальгетиками. Последние могут быть вытеснены их антагонистами, итогом чего становится восстановление болевой чувствительности. Налоксон блокирует, главным образом, опиатные рецепторы, в меньшей степени (в 10 раз) — о-опиатные рецепторы, и в наименьшей степени (в 30 раз) — к-опиатные рецепторы. Наряду с антагонистами опиоидных пептидов найдены также их агонисты.
Механизм обезболивающего действия опиоидных пептидов заключается в том, что после взаимодействия эндорфинов и энкефалинов с опиатными рецепторами не проявляется алгогенное действие субстанции Р и других алгогенов.
Механизм действия налоксона, имеющего меньший, чем опиоидные пептиды, размер молекулы, заключается в более быстрой и более сильной связи с опиатными рецепторами, в результате чего опиоиды не могут с ними взаимодействовать, а значит оказывать обезболивающее действие. 23.Серотонинэргическая система мозга. Серотонинергическая система осуществляет связь гипоталамуса с отделами среднего и продолговатого мозга и лимбической системы. Серотонинергические волокна поступают в срединное возвышение и заканчиваются в его капиллярах. Серотонин ингибирует гонадотропинрегулирующую функцию гипоталамуса на уровне аркуатных ядер.
Не исключено его опосредованное влияние через эпифиз.
Кроме биогенных аминов, в качестве нейромедиаторов, регулирующих гонадотропинрегулирующую функцию гипоталамуса, могут выступать опиоидные пептиды — вещества белковой природы, обладающие морфиноподобным действием. К ним относятся метионин- и лейцин-энкефалины, α-, β-, γ-эндорфины.
Основная масса опиоидов представлена энкефалинами. Они обнаружены во всех отделах ЦНС. Опиоиды изменяют содержание биогенных аминов в гипоталамусе, конкурируя с ними за рецепторные места [Бабичев В. Н., Игнатков В. Я-, 1980; Klee N., 1977]. Опиоиды оказывают ингибирующее воздействие на гонадотропную функцию гипоталамуса.
Роль нейротрансмиттеров и нейромодуляторов в ЦНС могут исполнять различные нейропептиды, найденные в большом количестве в различных отделах ЦНС. К ним относятся нейротензин, гистамин, субстанция Р, холецистокинин, вазоактивный кишечный пептид. Эти вещества оказывают преимущественно ингибирующее воздействие на продукцию люлиберина. Синтез гонадотропин-рилизинг-гормона (ГТ-РГ) стимулируют простагландины из группы Е и F2a [Ojeda S., McCann S., 1975].
Эпифиз — шишковидная железа — расположен в каудальной части III желудочка. Эпифиз имеет дольчатое строение и разделяется на паренхиму и соединительнотканную строму.
представлена клетками двух типов: пинеальными и глиальными. С возрастом количество клеток паренхимы уменьшается, увеличивается стромальная прослойка. К 8—9 годам в эпифизе появляются очаги обызвествления. Возрастную эволюцию претерпевает и сосудистая сеть, питающая эпифиз. Вопрос об инкреторной функции эпифиза остается нерешенным.
Из веществ, обнаруженных в эпифизе, наибольший интерес в плане регуляции гонадотропной функции представляют индольные соединения — мелатонин и серотонин. Эпифиз считают единственным местом синтеза мелатонина — производного серотонина, так как только в эпифизе найден специфический фермент гидроксииндоло-метил-трансфераза, осуществляющий конечный этап его образования.
Ингибирующее влияние эпифиза на половую функцию доказано в многочисленных экспериментальных работах. Предполагается, что свою антигонадотропную функцию мелатонин реализует на уровне гипоталамуса, блокируя синтез и секрецию люлиберина. Кроме того, в эпифизе обнаружены и другие вещества пептидной природы с выраженным антигонадотропным действием, превышающим активность мелатонина в 60—70 раз [Benson S., 1972]. Функция эпифиза зависит от освещенности. В связи с этим не исключена роль эпифиза в регуляции суточных ритмов организма, в первую очередь ритмов тропных гормонов гипофиза. 24.Возбуждающие медиаторы-аминокислоты. Типы рецепторов к глутамату и кратковременная память. Глутаминовая кислота (глутамат) – это главный возбуждающий медиатор ЦНС. Будучи заменимой пищевой аминокислотой, она широко распространена в самых разных белках, и ее суточное потребление составляет не менее 5-10 г. Однако глутаминовая кислота пищевого происхождения в норме очень плохо проникает через гемато-энцефалический барьер, что предохраняет от серьезных сбоев в деятельности мозга. Практически весь необходимый ЦНС глутамат синтезируется прямо в нервной ткани. Данное вещество является также промежуточной стадией в процессах внутриклеточного обмена аминокислот. Поэтому нервные клетки содержат довольно много глутаминовой кислоты, лишь небольшая часть которой выполняет собственно медиаторные функции. Синтез такого глутамата происходит прямо в пресинаптических окончаниях; основной предшественник – аминокислота глутамин. Выделяясь в синаптическую щель, медиатор действует на соответствующие рецепторы. Разнообразие рецепторов к глутаминовой кислоте чрезвычайно велико. В настоящее время выделяют три типа ионотропных и до восьми типов метаботропных рецепторов. Последние менее распространены и менее изучены. Их эффекты могут реализоваться как путем подавления активности аценилатциклазы, так и через усиление образования диацилглицерола и инозитолтрифосфата.
Аспарагиновая кислота (аспартат) также может выполнять в ЦНС функции возбуждающего медиатора. По своей химической формуле она очень близка к глутаминовой и действует на те же рецепторы. Эта кислота похожа на глутаминовую и действует на те же рецепторы. Встречается этот медиатор относительно редко. Так, в спинном мозгу аспартат содержится в возбуждающих интернейронах, регулирующих различные врожденные рефлексы. Много аспартата в нижней оливе – особом ядре на вентральной (передней) поверхности продолговатого мозга. Именно он является медиатором лазающих волокон, направляющихся из нижней оливы к мозжечку. Входя в кору мозжечка лазающие волокна образуют синапсы на клетках Пуркинье. Срабатывание таких синапсов воздействует на системы вторичных посредников и вызывает различные метаболические изменения. В результате на длительное время (несколько часов) снижается эффективность синапсов между параллельными волокнами и дендритами клеток Пуркинье. Это явление названо долговременной депрессией. Оно играет важную роль в процессах моторного обучения. При повреждении нижней оливы выработка новых двигательных навыков резко затрудняется. 25.Тормозные медиаторы-аминокислоты. ГАМК является непищевой аминокислотой. Это значит, что она не входит в состав белков и полностью синтезируется в нашем организме. ГАМК присутствует в нервной системе в большом количестве. Дело в том, что она, как и глутаминовая кислота, играет важную роль в процессах внутриклеточного обмена веществ (прежде всего, в ферментативном разложении глюкозы). И лишь небольшая часть ГАМК выполняет функции медиатора. В этом случае она легко образуется из глутаминовой кислоты прямо в пресинаптических окончаниях. Далее происходит перенос ГАМК в везикулы и выброс в синаптическую щель. ГАМК очень широко распространена в ЦНС – не менее широко, чем глутаминовая кислота. В основном она является медиатором относительно мелких нейронов, осуществляющих тормозную регуляцию проведения сигналов. Иными словами, передачу информации от одной нервной структуры к другой осуществляют, прежде всего, глутаматергические нейроны (релейные, тип Гольджи I). Функции же возвратного, латерального и прочего торможения реализуются преимущественно за счет деятельности ГАМК-ергических клеток. Однако, в некоторых зонах ЦНС встречаются и крупные релейные нейроны, использующие ГАМК в качестве медиатора. Таковыми являются, например, клетки Пуркинье (кора мозжечка) и клетки бледного шара, играющие чрезвычайно важную роль в двигательных центрах головного мозга. Выделяясь в синаптическую щель, ГАМК действует на соответствующие рецепторы. Выделяют два их типа – ГАМКА и ГАМКВ. Первый является постсинаптическим, ионотропным и содержит Cl--каналы; второй – как пост-, так и пресинаптическим, метаботропным и влияет на К+-каналы. Более исследованы ГАМКА-рецепторы, агонисты которых нашли широкое применение в клинической практике. Антагонисты ГАМКА-рецепторов бикукулин и пикротоксин являются сильными ядами и вызывают судороги. При этом бикукулин является конкурентным антагонистом и связывается с местом присоединения к рецептору самой ГАМК. Пикротоксин – неконкурентный антагонист и блокирует хлорный ионный канал. Глицин - заменимая пищевая аминокислота. Одновременно он является тормозным медиатором, хотя и значительно менее распространенным, чем ГАМК. Большая часть глицинергических клеток выполняет так называемое возвратное торможение. Его назначение - предохранение мотонейронов от перевозбуждения. Оно осуществляется следующим образом (рис. 14). Коллатераль от аксона мотонейрона образует возбуждающий синапс на интернейроне, который называется клеткой Реншоу (по имени исследователя, открывшего возвратное торможение). Аксон же клетки Реншоу направляется назад к мотонейрону и образует на нем тормозные синапсы. При слабом возбуждении мотонейрона, сопровождаемом одиночными ПД, бегущими по его аксону, возбуждение клетки Реншоу недостаточно для генерации ею нервного импульса. Однако когда возбуждение мотонейрона нарастает, частота ПД, проводимых по его аксону увеличивается. Это приводит к суммации ВПСП на клетке Реншоу и генерации ею серии импульсов, которые в конечном итоге притормаживают мотонейрон. 26. Электрические и химические синапсы: их строение и функции. СИ́НАПС (греч. synapsis — соединение, связь), зона контакта между нейронами и другими образованиями (нервными, мышечными или железистыми клетками), служащая для передачи информации от клетки, генерирующей нервный импульс к другим клеткам. Термин ввел Ч. Шеррингтон в 1897.
Синапс состоит из трех отделов: пресинаптического (нейрон, посылающий сигналы), постсинаптического (клетка, принимающая сигналы) и соединяющей их структуры (синаптическая щель). В тех случаях, когда речь идет о контактах между нервными клетками, синапсы могут образовываться между аксонамии сомой, аксонами и дендритами, аксонами и аксонами, дендритами и дендритами, а также между сомой и дендритами нейронов. В зависимости от способа передачи возбуждения выделяют химические (наиболее распространенные) и электрические синапсы. Существуют также смешанные синапсы, сочетающие оба механизма передачи.
Электрические синапсы распространены у беспозвоночных и низших позвоночных, но иногда встречаются и в некоторых участках мозга млекопитающих. Они образуются чаще всего между дендритами близко расположенных нейронов и осуществляют быструю (без синаптической задержки) передачу сигналов, благодаря наличию высокопроводящего контакта, обусловленного наличием узкой синаптической щели и специальных ультраструктур, снижающих электрическое сопротивление в области контакта.
Химические синапсы преобладают в мозгу млекопитающих. На соме и дендритах каждого нейрона может локализоваться до нескольких десятков тысяч синаптических окончаний. В их пресинаптических окончаниях содержатся синаптические пузырьки (везикулы), содержащие химический посредник, называемый медиатором (нейромедиатор, нейротрансмиттер) и имеющие различные размеры и электронную плотность. Так, обнаружены малые прозрачные пузырьки, заполненные низкомолекулярными, так называемыми, «классическими» медиаторами (ацетилхолин, ГАМК, глицин и др.) и крупные электронно-плотные, содержащие пептидные медиаторы. Медиаторы образуются в соме нейрона и затем по аксону транспортируются в синаптическое окончание. Согласно сформулированному в 1930-х годах закону Дейла, медиатор, обнаруженный в одном синапсе, должен быть также медиатором во всех других синаптических окончаниях того же нейрона. Позже выяснилось, что в одном нейроне может синтезироваться и в одном окончании освобождаться более одного медиатора, однако набор медиаторов для данного нейрона всегда постоянен.
Приходящий электрический импульс при участии ионов кальция вызывает освобождение медиатора из пресинаптических окончаний. Медиатор диффундирует через синаптическую щель шириной 10 — 50 нм и взаимодействует с рецепторными белками постсинаптической мембраны, что приводит к возникновению постсинаптического потенциала. Время, в течение которого происходят эти реакции, называется синаптической задержкой и составляет 0,3 — 1 мс. Не связавшийся с рецептором медиатор либо разрушается специальными ферментами, либо захватывается обратно в пузырьки пресинаптического окончания.
Характерным для синапсов является их особенность изменять чувствительность к действию медиаторов в процессе своей активности. Это свойство называется синаптической пластичностью и составляет основу таких процессов, как память и обучение. Различают кратковременную синаптическую пластичность, продолжающуюся не более 20 мин, и долговременную, длящуюся от нескольких десятков минут до нескольких недель. Пластичность может проявляться как в форме потенциации (активации), так и в форме депрессии. В ее основе лежат различные механизмы от изменения концентрации ионов кальция в синаптической области до фосфорилирования или разрушения синаптических белков, а также экспрессии или репрессии генов, катализирующих синтез таких белков. В зависимости от степени пластичности синапсы разделяют на стабильные и динамические, причем первые формируются в онтогенезе раньше, чем последние. 27.Процессы, происходящие в нервно-мышечном синапсе. Нервно-мышечный синапс (также нейромышечный, либо мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне. Входит в состав нервно-мышечного веретена. Нейромедиатором в этом синапсе является ацетилхолин.
В этом синапсе нервный импульс превращается в механическое движение мышечной ткани.
Волокна скелетных мышц иннервируются аксонами нервных клеток, называемых мотонейронами (или соматическими эфферентными нейронами) .
Аксоны мотонейронов , расположенных в передних рогах спинного мозга ( двигательные аксоны ) образуют синапсы с волокнами скелетных мышц .
При подходе аксона к поверхности мышечного волокна миелиновая оболочка заканчивается, и он образует терминальную часть (нервное окончание) в виде нескольких коротких отростков, располагающихся в желобках на поверхности мышечного волокна. Область плазматической мембраны мышечного волокна, лежащая непосредственно под нервным окончанием, обладает особыми свойствами и называется двигательной концевой пластинкой. Структура, состоящая из нервного окончания и двигательной концевой пластинки, - это нервно-мышечное соединение (нервно-мышечный синапс) Таким образом, двигательной концевой пластинкой (нервно-мышечным соединением, нервно-мышечными концевыми пластинками, моторными бляшками) называются синапсы между аксоном мотонейрона и волокном скелетной мышцы.
Они имеют все типичные морфологические характеристики химических синапсов.
Рассмотрим нервно-мышечное соединение скелетной мышцы при возбуждении мембраны мышечного волокна .
Поскольку сигналом для запуска сокращения служит потенциал действия плазматической мембраны волокна скелетной мышцы резонно задать вопрос: каким образом он возникает? В скелетных мышцах потенциалы действия можно вызвать только одним способом - раздражением нервных волокон . (Для инициации сокращений сердечной мышцы и гладкой мускулатуры есть и другие механизмы).
Итак, как было сказано выше, волокна скелетных мышц иннервируются аксонами нервных клеток (мотонейронами). Тела этих клеток расположены в стволе мозга или в спинном мозге . Аксоны мотонейронов покрыты миелиновой оболочкой, а их диаметр больше, чем у других аксонов, поэтому они проводят потенциалы действия с высокой скоростью, обеспечивая поступление сигналов из ЦНС к волокнам скелетных мышц лишь с минимальной задержкой.
При подходе аксона к поверхности мышечного волокна миелиновая оболочка заканчивается, и он образует терминальную часть (нервное окончание) в виде нескольких коротких отростков, располагающихся в желобках на поверхности мышечного волокна (аксон мотонейрона разделяется на множество ветвей, каждая из которых образует одно соединение с мышечным волокном) . Таким образом, один мотонейрон иннервирует много мышечных волокон, но каждым мышечным волокном управляет ветвь только от одного мотонейрона. Область плазматической мембраны мышечного волокна, лежащая непосредственно под нервным окончанием, обладает особыми свойствами и называется двигательной концевой пластинкой , а мотонейрон и мышечные волокна, которые он иннервирует, составляют двигательную единицу . Мышечные волокна одной двигательной единицы находятся в одной и той же мышце, но не в виде компактной группы, а рассеяны по нем.. Когда в мотонейроне возникает потенциал действия, все они получают стимул к сокращению. Структура, состоящая из нервного окончания и двигательной концевой пластинки, - это нервно-мышечное соединение (нервно-мышечный синапс) 28.Постсинаптические потенциалы, их отличие от ПД. Суммация в ЦНС. Возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10—15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД. ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора. В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
Суммация - явление суммирования деполяризующих эффектов нескольких возбуждающих постсинаптических потенциалов, каждый из которых не может вызвать деполяризацию пороговой величины, необходимой для возникновения потенциала действия.
Потенциалы действия, генерируемые разными нейронами, примерно одинаковы, постсинаптические потенциалы, возникающие в разных входных синапсах на одном и том же нейроне, сильно варьируют как по величине, так и по продолжительности. В одном синапсе на мотонейроне приходящий нервный импульс может вызвать деполяризацию в 0,1 мВ, а в другом - деполяризацию в 20 мВ. Если степень деполяризации окажется одинаковой, эффект будет тем сильнее, чем больше область синаптического контакта, но природа системы такова, что даже малые по величине постсинаптические потенциалы, суммируясь, могут давать большой эффект.
Индивидуальные постсинаптические потенциалы, как правило, не приводят к возникновению потенциала действия. Если сигналы одновременно приходят к нескольким синапсам, находящимся в одном и том же участке дендрита, то общий постсинаптический потенциал будет примерно равен сумме отдельных постсинаптических потенциалов, причем тормозные постсинаптические потенциалы суммируются с отрицательным знаком. Суммарное электрическое возмущение, возникшее в одном постсинаптическом участке, будет распространяться на другие участки благодаря пассивным кабельным свойствам мембраны дендрита.
Благодаря временной суммации и пространственной суммации потенциалы действия множества нейронов могут определять мембранный потенциал одного постсинаптического нейрона, в результате чего выработается определенный ответ, обычно в виде импульсов для передачи сигналов другим клеткам. Ответный сигнал должен отражать величину суммарного постсинаптического потенциала, которая может плавно изменяться. Однако потенциалы действия имеют постоянную амплитуду и распространяются по закону "все или ничего". Единственной свободной переменной при передаче сигналов с помощью импульсов остается временной интервал между последовательными импульсами. Поэтому для передачи информации величина суммарного постсинаптического потенциала должна быть преобразована (перекодирована) в виде частоты импульсного разряда. Такое кодирование достигается с помощью специальной группы потенциал-зависимых ионных каналов, расположенных в основании аксона. 29.Локальные тормозные нейронные сети. Пресинаптическое и постсинаптическое торможение. Взаимодействующие между собой посредством передачи через отростки возбуждений нейроны формируют нейронные сети. Переход от рассмотрения отдельного нейрона к изучению нейронных сетей является естественным шагом в нейробиологической иерархии. Нейроны образуют два характерных типа соединений - конвергентные, когда большое число нейронов одного уровня контактирует с меньшим числом нейронов следующего уровня, и дивергентные, в которых контакты устанавливаются со все большим числом клеток последующих слоев иерархии. Сочетание конвергентных и дивергентных соединений обеспечивает многократное дублирование информационных путей, что является решающим фактором надежности нейронной сети. При гибели части клеток, сохранившиеся нейроны оказываются в состоянии поддерживать функционирование сети. Ко второму типу нейронных сетей относятся локальные сети, формируемые нейронами с ограниченными сферами влияния. Нейроны локальных сетей производят переработку информации в пределах одного уровня иерархии. При этом функционально локальная сеть представляет собой относительно изолированную тормозящую или возбуждающую структуру. Важную роль также играют так называемые дивергентные сети с одним входом. Командный нейрон, находящийся в основании такой сети может оказывать влияние сразу на множество нейронов, и поэтому сети с одним входом выступают согласующим элементом в сложном сочетании нейросетевых систем всех типов.
Пресинаптическое торможение - это уменьшение или прекращение высвобождения медиатора из пресинаптических нервных окончаний. При этом не происходит генерации тормозного постсинаптического потенциала.Преимущество пресинаптического торможения состоит в его избирательности, так как происходит торможение отдельных входов нервной клетки, в то время как при постсинаптическом торможении снижается возбудимость всего нейрона. Снижение количества высвобождающегося медиатора в случае пресинаптического торможения связано с активацией аксо- аксонных синапсов, и, вероятно, обусловлено снижением амплитуды пресинаптического потенциала действия в результате инактивации.
Постсинаптическое торможение - это снижение возбудимости постсинаптической мембраны нейрона, препятствующее распространению импульса. Нервный импульс в тормозных нейронах вызывает гиперполяризационный сдвиг потенциала, в результате чего уровень мембранного потенциала начинает сильнее отличаться от порогового потенциала , необходимого для генерации потенциала действия . Поэтому гиперполяризация постсинаптической мембраны называется тормозным постсинаптическим потенциалом . Механизм высвобождения медиатора в тормозных синапсах и возбуждающих синапсах , видимо, аналогичен. Тормозным медиатором в мотонейронах и некоторых других синапсах служит аминокислота глицин . Медиатор, действуя на постсинаптическую мембрану, открывает поры, или каналы, через которые могут проходить все мелкие ионы. Если стенка поры несет электрический заряд, то он препятствует прохождению одноименно заряженных ионов. При одновременном возникновении возбуждающих и тормозных синаптических процессов амплитуда возбуждающего постсинаптического потенциала уменьшается в зависимости от амплитуды тормозного постсинаптического потенциала . 30.Функции спинного мозга. Спинной мозг выполняет рефлекторную и проводниковую функции. Первая обеспечивается его нервными центрами, вторая проводящими путями. Он имеет сегментарное строение. Причем деление на сегменты является функциональным. Каждый сегмент образует передние и задние корешки. Задние являются чувствительными, т.е. афферентными, передние - двигательными, эфферентными. Эта закономерность называется законом Белла-Мажанди. Корешки каждого сегмента иннервируют 3 метамера тела, но в результате перекрывания каждый метамер иннервируется тремя сегментами. Поэтому при поражении передних корешков одного сегмента, двигательная активность соответствующего метамера лишь ослабляется.
Морфологически тела нейронов спинного мозга образуют его серое вещество. Функционально все его нейроны делятся на мотонейроны, вставочные, нейроны симпатического и парасимпатического отделов вегетативной, нервной системы. Мотонейроны, в зависимости от функционального значения, разделяются на альфа- и гамма-мотонейроны. К мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных. т.е. рецепторных мышечных клеток. Тела а-мотонейронов расположены в передних рогах спинного мозга, а та аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом, они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает.
Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС. Нейроны симпатического отдела вегетативной нервной системы находятся в боковых рогах грудных сегментов, а парасимпатического в крестцовом отделе.
Проводниковая функция сострит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные или проприоспинальные, восходящие и нисходящие. Проприоспинальные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации то
|