| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
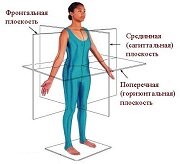
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| ФИЗИОЛОГИЯ ТРОЙНИЧНОГО НЕРВА
Ветви тройничного нерва осуществляют иннервацию различных тканей головы. Большинство рецепторов этой системы возбуждаются только при непосредственном контакте с раздражителем. Примером высокой специализации может служить фациальная ямка некоторых змей, с помощью которой эти животные эффективно локализуют на расстоянии источник теплового излучения. У млекопитающих телерецепция такого типа отсутствует, но у них хорошо развиты вибриссы, с помощью которых животные активно исследуют окружающую среду. Самой высокой сложности тригеминальная система достигает у человека в связи с ее участием в речевой функции. Все многочисленные ветви тройничного нерва объединяются в три подразделения: глазничное, максиллярное и мандибулярное (рис. 6.23). У входа в головной мозг они образуют крупный полулунный (гассеров) ганглий, образованный телами псевдоуниполярных нейронов. Зрительная ветвь иннервирует кожу лба и носа, роговицу, конъюнктиву и внутриглазные структуры, слизистую параназальных синусов, верхней, нижней и передней частей перегородки носа и латеральной стенки носовой полости, слезный проток. Верхнечелюстная ветвь иннервирует область кожи верхней челюсти, слизистую максиллярного синуса, верхние зубы и десны, слизистую задней части перегородки носа и нижней части носовой полости, верхние зубы и десны, твердое небо и миндалины. Нижнечелюстная ветвь иннервирует область кожи нижней челюсти, слизистую щек, дно носовой полости, язык, проприорецепторы мышц нижней челюсти, нижние зубы и десны, жевательные мышцы. Все подразделения тройничного нерва иннервируют мозговые оболочки и сосуды мозга.
Некоторая часть первичных афферентов зрительной и верхнечелюстной ветвей, а также значительное количество волокон мандибулярной ветви не имеют клеточных тел в гассеровом ганглии. Тела этих афферентных нейронов находятся в тройничном мезэнцефалическом ядре, которое представляет собой узкую полоску псевдоуниполярных нейронов, протянувшуюся вдоль всего среднего мозга. С филогенетической точки зрения тройничное мезэнцефалическое ядро, как и гассеров ганглий, является гомологом спинномозговых ганглиев. Аксоны нейронов этого ядра проходят с моторными ветвями тройничного нерва к жевательным мышцам и осуществляют афферентную иннервацию проприорецепторов (мышечных веретен и сухожильных рецепторов). Они также иннерви-руют рецепторы зубов, языка, сустава нижней челюсти; при этом большая часть их направляется ипсилатерально, но некоторые отсылают свои аксоны на контралатеральную сторону. Часть волокон мезэнцефалического ядра заканчивается в моторном тройничном ядре, образуя двухнейронную дугу жевательных рефлексов, но некоторая часть спускается к ядрам спинальных тригеминальных ядер, расположенных на уровне продолговатого мозга. Центральные отростки нейронов мезэнцефалического тригеминального ядра объединяются в латеральный продольный пучок, который, таким образом, является афферентной частью рефлекторных систем, осуществляющих контроль кусания, жевания, глотания, участвует в сокращении наружных глазных мышц и мимической мускулатуры. Афференты сенсорного корешка тройничного нерва в мозге делятся на толстые нисходящие и тонкие восходящие волокна. Восходящая часть волокон направляется к главному сенсорному ядру (на уровне моста), а более толстые нисходящие волокна образуют нисходящий спинальный тройничный тракт, который спускается вниз и достигает уровня первых шейных сегментов, синаптически контактируя с ядрами тройничного тракта. В составе тройничного тракта, кроме первичных афферентов, имеется значительное количество аксонов нейронов 2-го порядка. В составе волокон тройничного тракта имеются волокна лицевого, языкоглоточного и блуждающего нервов. В настоящее время различают два компонента тригеминального комплекса: главное сенсорное ядро (локализовано в мосте) и сенсорные ядра спинотригеминального тракта (локализованы в продолговатом мозге и верхних шейных сегментах). Филогенетические данные и характер центральных связей тройничных сенсорных ядер позволяют рассматривать главное сенсорное ядро как гомолог ядер дорсальных столбов спинного мозга, тогда как ядра тригеминального тракта как гомолог серого вещества дорсальных рогов спинного мозга. В составе тройничных нервов идут также волокна вегетативной нервной системы. Парасимпатические волокна берут начало от верхнего и нижнего слюнных ядер продолговатого мозга. Симпатические волокна являются аксонами нейронов боковых рогов спинного мозга. Вегетативные волокна, входящие в состав ветвей тройничного нерва, регулируют секрецию желез слизистой оболочки рта и носа. Функциональные характеристики афферентных волокон тройничного нерва. Афферентные волокна тройничного нерва проводят афферентную импульсацию разной модальности - от механорецепторов, терморецепторов и болевых рецепторов. Окончания тройничного нерва образуют два типа терморецепторов: специфические терморецепторы реагируют только на изменение температуры, неспецифические - как на температурные, так и на механические воздействия. Терморецепторы у большинства млекопитающих локализованы в структурах ротовой полости и кожи носа. Пульпа зубов содержит только болевые рецепторы, которые иннервируются толстыми волокнами. Механорецепторы, от которых идет импульсация по волокнам тройничного нерва, тонко специализированы, что обеспечивает точность оценки механического стимула в области морды животного. Особенно тонкий анализ механических стимулов у млекопитающих осуществляется рецепторами волос. У них выделяют три типа волос: вибриссы (сенсорные волосы), остевые волосы (тилотрихи) и волосы шерсти. Каждому типу волос соответствуют особенности строения волосяных луковиц. Особенно велико значение вибрисс. У крысы, например, одну луковицу вибриссы иннервируют 100-150 нейронов гассерова ганглия, при этом отдельные нейроны передают в мозг информацию о таких признаках, как исходное положение вибриссы, направление, скорость и амплитуда ее отклонения и т. д. Афферентные волокна, передающие импульсы от механорецепторов в области морды, имеют очень маленькие рецептивные поля по сравнению с афферентными волокнами, иннервирующими поверхность лап или туловища. Например, рецептивные поля на морде кошки имеют размер 7 - 15 мм2, а вибрисс - 1 мм2. На гладкой коже носа у кошки, собаки и обезьяны размер рецептивного поля - тоже порядка 1 мм2. Рецепторы растяжения наружных мышц глаза у млекопитающих иннервируются тонкими аксонами нейронов, тела которых находятся в мезэнцефалическом ядре (кошка, человек), но у некоторых животных (овца, свинья) - в гассеровом ганглии. Проекция первичных афферентов в ядра тройничного нерва исключительно ипсилатеральная и организована в виде «листков», которые идут в рострокаудальном направлении, включая по нескольку ядер комплекса. Нейроны, получающие сигналы из полости рта, локализованы в более медиальных областях тройничных сенсорных ядер, тогда как периоральные структуры и кожа головы представлены в них латерально. У млекопитающих самую обширную группу в тройничных сенсорных ядрах образуют нейроны, связанные с тактильными рецепторами, особенно те, которые возбуждаются при сгибании вибрисс; эти нейроны занимают в ядрах дорсолатеральную часть. У опоссума, например, нейроны, связанные с тактильными рецепторами вибрисс и гладкой кожи носа, занимают 4/5 объема тройничных сенсорных ядер. Центральную часть тройничных сенсорных ядер занимает столб клеток, на которые проецируются рецепторы зубов. Она простирается от центральной области главного сенсорного ядра до рострального ядра спинального тригеминального тракта. Нейроны, связанные со специфическими терморецепторами языка, локализуются в каудальном ядре спинального тригеминального тракта. Нейроны, которые активируются от неспецифических терморецепторов кожи, обнаружены как в каудальном, так и в оральном ядрах спинального тригеминального тракта. В главном сенсорном ядре, в оральном и каудальном ядрах преобладают нейроны с локальными рецептивными полями, тогда как нейроны интерполярного ядра имеют преимущественно обширные рецептивные поля. В дорсальной части орального ядра обнаружено большое количество нейронов, на которых заканчиваются афференты от периоральной области и вибрисс. Тройничное моторное ядро, локализованное вентральнее сенсорного ядра, состоит из большого дорсального подразделения, образованного редко расположенными крупными нейронами с хорошо развитыми дендритами. Вентральная часть ядра образована более мелкими, но более тесно расположенными нейронами. Нейроны этого ядра – мотонейроны - осуществляют эфферентную иннервацию мышечных волокон жевательных мышц. В ядре существует соматотопия: мышцы верхней части головы (жевательная и височная) иннервируются мотонейронами, расположенными в вентральном подразделении ядра, тогда как мышцы нижней части головы (наружная и внутренняя крыловидная мышцы) - нейронами дорсальных отделов ядра. Полная дифференцировка ядер тригеминальной системы заканчивается только у млекопитающих. Связи тригеминальной системы с таламусом и мозжечком. Почти все нейроны главного тригеминального сенсорного ядра посылают свои аксоны к таламусу (см. рис. 6.23). Нейроны, аксоны которых формируют билатеральные лемнисковые тракты, составляют 2/3 вентральной части главного сенсорного ядра. Ипсилатеральный тригеминальный тракт образован аксонами нейронов, тела которых локализованы в дорсальной части ядра. От ядер спинального тройничного тракта берут начало два тригеминоталамических пути -вентральный и дорсальный. Вентральный тригеминальный тракт начинается от каудального ядра, затем переходит на контралатеральную сторону и заканчивается в заднемедиальном вентральном ядре и частично в интраламинарных ядрах таламуса. Дорсальный тригеминальный тракт начинается от нейронов каудального ядра. Это билатеральный тракт, но с преобладанием контралатеральной компоненты. Достигнув таламуса, волокна тракта оканчиваются в заднемедиальном вентральном и интраламинарных ядрах таламуса. Мозжечок получает проекции от нейронов тригеминальных ядер. Основным источником этих волокон являются ядра спинального тригеминального тракта. Их аксоны оканчиваются преимущественно рострально в коре червя мозжечка. Таким образом, тригеминоталамические пути распределены в структурах как специфического, так и неспецифического отделов таламокортикальной системы. Нейроны главного сенсорного тригеминального ядра не имеют проекций на ретикулярную формацию, тогда как ядра спинального тригеминального тракта образуют как ипси-, так и контралатеральные проекции на парамедианное и гигантоклеточное ядра ретикулярной формации. Таламический и корковый уровни анализа кожно-кинестетической информации. Ядра вентробазального таламуса переключают информацию о прикосновении, давлении, движении в суставах. Обработка этой информации на уровне специфических таламических ядер организована по принципу соматотопии и модальности. В отличие от этого для нейронов интраламинарных ядер, заднего и ретикулярного таламических ядер, к которым подходят волокна спиноталамических трактов, характерна обширная конвергенция ряда модальностей, включая болевую. Соматосенсорная кора имеет два подразделения: SI (первичная соматосенсорная область) и SII (вторичная соматосенсорная область). SI у приматов занимает переднюю часть теменной доли, включая дно и заднюю стенку центральной борозды. Другая, менее изученная, область - SII - лежит на передней стенке сильвиевой извилины. Кроме этого, соматосенсорные сигналы достигают нейронов моторной прецентральной коры (поля 4, 6) и поле 5 (ростральная теменная кора). Области SI и SII тесно связаны между собой. Поле 5 получает основной вход от латерального заднего таламического ядра, а также пространственно организованный вход от полей 1 и 2. Известно также, что симметричные зоны коры, включая области SI и SII, связаны между собой через мозолистое тело. В области SI коры выделяют несколько зон, соответствующих полям За, Зb, 1 и 2 (по Бродману). Нейроны поля За получают информацию от мышечных и суставных рецепторов, нейроны поля 2 - от глубоких структур (возбуждаются при сдавливании кожи и практически не реагируют на легкое прикосновение). В отличие от этого нейроны поля Зb реагируют на легкое прикосновение к коже, сгибание отдельных волосков. Нейроны поля 1 реагируют как на кожную стимуляцию, так и на стимуляцию глубоких структур. Характерным для сенсорных и моторных полей коры является организация нейронов, участвующих в обработке однородной информации, в вертикальные колонки. Например, нейроны одной колонки области SI реагируют на стимулы одной модальности. Эффекты разрушения сенсорных полей коры. Избирательное разрушение поля 3 приводит к тому, что, например, обезьяна на ощупь не способна отличать мягкий предмет от твердого, шероховатую поверхность от гладкой, выпуклую от вогнутой. Разрушение поля 2 нарушает способность различать форму у предметов. Например, невозможно было обучить обезьяну отличать при ощупывании квадрат от ромба. Разрушение поля 1 вызывает достаточно легкий дефицит в дискриминации рельефа поверхности при ощупывании. Было сделано заключение, что повреждение SI (в состав которой входят указанные поля) приводит не только к дефициту пространственного различения (например, определение величины углов, характера поверхности- вогнутая или выпуклая и др.), но и к дискриминации временных событий. Например, обезьяна была не способна различать осцилляции различной частоты, приложенные к поверхности кожи. Основные результаты экспериментальных исследований были подтверждены клиническими наблюдениями больных с поврежденной постцентральной областью коры. У таких больных нарушалась дискриминация размера и формы предметов. Например, больные не могли отличить на ощупь овал от параллелепипеда (в клинике это нарушение носит название астерогнозис). Как у человека, так и у обезьяны при повреждении постцентральной коры страдает кинестезия, что проявляется в резком ухудшении способности оценивать положение конечности при пассивном или активном ее перемещении. Страдает также оценка силы, приложенной к руке.Ощущение боли и температуры при этих повреждениях коры сохраняется. При поражении вторичных отделов постцентральной коры у больных возникают своеобразные явления «афферентной апраксии». Они проявляются в том, что больной не способен выполнять тонкие дифференцированные движения. При поражении аналогичных зон левого (доминантного) полушария с распространением на нижние отделы (область представительства лица, губ и языка) кинестетическая апраксия может проявляться в нарушении движений речевого аппарата (афферентная моторная афазия). Нарушения проявляются в невозможности найти положение губ и языка, необходимые для артикуляции соответствующих звуков. Больные смешивают близкие артикулемы, например заменяют «б» или «п» на «м». Другими словами, больные смешивают акустически разные, но близкие по артикуляции буквы. При этих поражениях коры страдает также и письмо. Больной может вместо слова «халат» написать «хадат» или «ханат». Считают, что такие симптомы являются результатом нарушения афферентной основы движения. Вопросы 1. Модальности соматической системы: механорецепция, терморецепция, боль, проприорецепция. 2. Сенсорные (восходящие) пути спинного мозга; нейрофизиологическая характеристика. 3. Иннервация головы - система тройничного нерва. Литература Основы сенсорной физиологии./Под ред. Р. Шмидта. М.: Мир, 1984.
|