| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
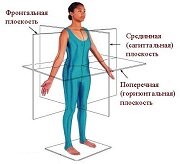
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Носители для иммобилизации ферментов
Содержание Инженерная инзимология Введение ГЛАВА 1. Иммобилизованные ферменты ГЛАВА 2. Иммобилизованные клетки
Введение Инженерная энзимология изучает био каталитические процессы в различных отраслях народного хозяйства, решая задачи получения и применения ферментов (энзимов). Эти процессы совершают особые белки — биологические катализаторы, ускоряющие химические реакции. Инженерная энзимология - это новое научно-техническое направление, появление которого связано с необходимостью интенсификации биотехнологических процессов. Задачи инженерной энзимологии - конструирование органических катализаторов с заданными свойствами на основе ферментов и полиферментных систем, выделенных из клеток или находящихся в них. Термин "заданные свойства" имеет в виду, что эти свойства задаются практическими потребностями в данном катализаторе, условиями проведения ферментативного процесса, специфичностью, необходимой производительностью и т.д. Инженерная энзимология основана на принципах органического и ферментативного катализа, химической технологии, биотехнологии и биохимии и со времени своего зарождения целиком обращена к практике. Хотя основная направленность современной инженерной энзимологии - использование каталитической активности иммобилизованных ферментов и клеток, ее рамки гораздо шире. Цель этой дисциплины - разработка научных основ применения ферментных катализаторов для создания новых биотехнологических производств, новых методов в диагностике и терапии, органическом синтезе и др., а также решение фундаментальных проблем энзимологии при помощи иммобилизованных ферментов. Практические разработки в области инженерной энзимологии связаны с : § получением нового продукта; § улучшением качества известного продукта; § повышением экономичности биотехнологического процесса.
Глава 1 Первые систематические исследования специфических ферментов проведены Эмилем Фишером в начале XX в. К тому времени уже было известно, что ферменты — наиболее специализированные белки, обладающие каталитической активностью. Их эффективность и мощность намного превосходят синтетические субстратам и ускоряют строго определенные химические реакции без образования побочных продуктов; ферменты функционируют температуры и рН. Ферменты в качестве биологических катализаторов применяются в различных отраслях промышленности - пищевой, текстильной, фармацевтической, кожевенной, в медицине, сельском хозяйстве, в тонком органическом синтезе и т.д. Более широкое использование ферментов в биотехнологии до последнего времени сдерживалось вследствие: Создание биокатализаторов нового поколения - иммобилизованных, т.е. связанных ферментов открыло перед прикладной энзимологией новые перспективы. Применение прикрепленных ферментов известно давно. Так, Дж.Нельсон и Е.Гриффин еще в 1916 г. показали, что инвертаза, адсорбированная на угле, сохраняет каталитическую активность. Но первый патент на применение иммобилизованных ферментов был выдан лишь в 1939 г. Дж.Пфанмюлеру и Г.Шлейху, предложившим для обработки шкур использовать протеолитические ферменты, адсорбированные на древесных опилках. В 1959 г. Н.Грубкофер и Д.Шлейт впервые применили для иммобилизации ферментов принципиально новый методический прием - ковалентное связывание. С этого времени и ведется целенаправленная разработка гетерогенных катализаторов на основе ферментов. В 1971 г. на первой конференции по инженерной энзимологии в Хенникере (США) был узаконен термин "иммобилизованные ферменты". Ферменты — функциональные единицы клеточного метаболизма. Действуя в строго определенной последовательности, они катализируют сотни многостадийных реакции, в ходе которых расщепляются молекулы питательных веществ запасается и преобразуется химическая энергия и из простых молекул-предшественников строятся макромолекулы входящие в состав клетки. С помощью ферментов обеспечивается гармоничное равновесие между различными метаболическими процессами, необходимыми для поддержания жизнеспособности отдельных клеток и организма в целом. В настоящее время известно и изучено почта 2000 различных ферментов, многие из которых успешно применяются и медицине, пищевой технологии, перерабатывающей промышленности, бытовой химии и других отраслях народного хозяйства. Основное внимание в ранний период развития энзимологии было сосредоточено на исследовании ферментов пищеварения пепсин, трипсин, химотринсин) и брожения, поэтому именно они наиболее изучены.Сейчас осознана важная роль внутриклеточных ферментов. Современный уровень развития энзимологии характеризуется открытием их новых представителей, которое позволило понять механизм многих фундаментальных жизненных процессов. Знания о закономерностях развития автолитических процессов, например при переработке промышленных животных, значительно пополнились благодаря выделению и изучению свойств внутриклеточных протеиназ (катепсинов) и ферментов, ответственных за двигательную функцию клетки. Насущной проблемой энзимологии является расшифровка механизма действия ферментов на основе изучения их химического строения, которое обеспечивает исключительно высокую и специфическую каталитическую активность. По химическому составу различают однокомпонентные ферменты - простые белки, сходные с глобулинами, которые при Гидролизе образуют только аминокислоты, и двухкомпонентные, представляющие собой сложные белки, в которых небелковым компонентом являются нуклеотиды, витамины, металлы и т. д. Именно благодаря ферментам клеток живого организма осуществляются различные химические превращения, обеспечивающие все процессы жизнедеятельности. Свое название ферменты получили от латинского слова «fermentum», что означает закваска, перебродивший сахар. Это соответствует первым представлениям о ферментах, которые были получены при изучении брожения (fermentation означает брожение). Синоним термина «ферменты» — энзимы происходит от греческого слова «enzyme», что означает закваска, дрожжи, с помощью которых начали изучать спиртовое брожение. Важная информация о природе участвующих в катализе химических групп молекулы фермента была получена при исследовании уникальной специфичности ферментов, которая не свойственна другим катализаторам, и при изучении действия ингибиторов и реагентов, атакующих определенные химические группы. Возможность получения ферментов в чистом виде привела к успешному развитию структурной энзимологии. Исследование чистых ферментов с помощью специальных химических методов позволило определить полные последовательности аминокислот в полипептидных цепях многих ферментов, а применение методов рентгеноструктурного анализа - установить детальную трехмерную структуру молекул некоторых ферментов, полученных в кристаллическом состоянии. Были определены конформация пептидных цепей, расположение различных групп, образующих субстратсвязывающий участок, и характер взаимодействия последнего с молекулой субстрата. Структура ферментов. На основе фундаментальных исследований установлено, что все ферменты обладают свойствами белков, причем их каталитическая активность зависит от степени сохранности нативнои структуры белка. Имея большую молекулярную массу и являясь полимерами, молекулы ферментов обладают компактной структурой отличающейся от структуры обычных линейных полимеров. Молекулы ферментов небольшие плотно «уложены», содержат всего лишь 20-30 % воды, их раствор имеют во много раз меньшую вязкость, чем растворы линейных полимеров. Первичная структура ферментов, как известно, представлена последовательностью аминокислот, соединенных в цепь пептид иными связями. Первичная структура расшифрована для таких протеолитических ферментов, как трипсиноген, химотрипсиноген. папаин и некоторых микробных протеиназ. Первичная структура известных ферментов представляет собой совокупность аминокислот, соединенных пептидными связями, по природе ковалентными и составляющими энергетический остов молекулы. Они характеризуются наибольшей электронной плотностью и связывают атомы, находящиеся на близком (0,1 — 0,2 нм) расстоянии. Вторичная структура молекул ферментов регулирует конформацию ее главной цепи. Она возникает в результате образования упорядоченной структуры в виде спирали посредством водородных связей между группами СО— и NH— и встречается в пептидных цепях на отдельных участках. Наиболее удачной моделью такой структуры признана модель Полинга и Кори, названная ими а-спиралью. Последняя не является единственным элементом вторичной структуры, в полипептидных цепях молекулы ферментов могут находиться также изгибы и повороты, нарушающие последовательность спирали. Встречаются также (3-складчатые структуры с вытянутыми цепями и межмолекулярными водородными связями. Вторичный уровень структуры поддерживается водородными (между электроотрицательными атомами через атом водорода) и электростатическими (типа солевых мостиков СОО — NH3 ) взаимодействиями. Под третичной структурой фермента понимают специфическую укладку регулярных и аморфных участков полипептидных цепей в глобулу. При этом углеводородные радикалы, направленные наружу по отношению к оси спирали, образуют гидрофобное ядро молекулы фермента. В создании и стабилизации третичной структуры принимают участие все типы химических связей, включая ковалентные. Имеются сведения, что в формировании третичной структуры нередко важную роль играют катионы металлов, например в а-амилазах это ионы кальция. Связывание отдельных участков полипептидной цепи металлом может быть довольно прочным, если мостик , образованный между металлом и специфической реагирующей группой, находится в глубинных слоях молекулы и, следовательно, защищен от воздействия молекул воды. Следует отметить особую роль гидрофобных взаимодействий (индуцированных и дисперсионных), прежде всего ван-дер-ваальсовых, а также образованных атомами молекулы воды. Поскольку аминокислотный состав ферментов весьма разнообразен, то и типы взаимодействий выражаются по-разному: для одних в совокупность взаимодействий основной вклад будут вносить электростатические, для других- гидрофобные, для третьих - ковалентные связи. В связи с этим каждый конкретный фермент будет иметь свои особый путь формирования белковой глобулы, который определяет специфические особенности и конформацию, в том числе при воздействии того или иного физико-химического фактора среды. Для некоторых ферментов установлена четвертичная структура, которая возникает в результате ассоциации двух или более глобулярных субъединиц с образованием составной или комплексной глобулы (например, папаин или а-амилаза Bacillus subtilis). При этом диссоциация или ассоциация составных глобул глубоко затрагивает активность ферментов. Следует, однако, отметить, что для большинства ферментов в сохранении каталитической активности решающую роль играет третичная структура. Ферментативный катализ происходит на расстоянии длины химической связи, поэтому катализ совершается на определенном участке поверхности макромолекулы — на активном центре. В результате экспериментальных исследований установлено, что активный центр представляет собой набор небольшого числа функциональных групп, расположенных близко друг от друга, и имеет вид впадин, выемок на поверхности молекулы фермента. Функциональные группы активного центра могут принадлежать звеньям полипептидной цепи, весьма удаленным друг от друга. Сближение их связано с формированием третичной структуры молекулы фермента. Ферментативный катализ имеет ряд особенностей. I. Ферменты проявляют высокую активность (активность — это количество молей субстрата, т. е. вещества, на которое действует фермент, превращаемое в продукт реакции одним молем фермента за 1 мин при оптимальных значениях рН и температуры), катализируя превращение субстратов при очень малых кон- центрациях. 2. Для ферментов характерна лабильность (чувствительность) к 'внешним факторам, к числу которых прежде всего относятся рН и температура. Так, у растений и у холоднокровных животных заметная активность ферментов начинает проявляться лишь при повышении температуры окружающей среды (весной), при низкой температуре (около 0С) ферменты почти не действуют. Для большинства ферментов наиболее благоприятной температурой является 37-40 С. при повышении температуры на 10 С активность ферментов приблизительно удваивается. Однако при увеличении температуры свыше 50 С происходит снижение активности ферментов, а при 80-100С она полностью утрачивается. Зависимость активности ферментов от рН носит аналогичный характер. Наилучшей, или оптимальной, средой для действия ферментов является слабокислая или слабощелочная среда. Изменение реакции среды в кислую или щелочную сторону ведет к снижению активности, а впоследствии к полной инактивации Каждый фермент проявляет свою активность при оптимальном значении рН. Вполне логично, что активный центр не имеет автономного существования, нельзя провести какую-либо определенную грань между активным центром фермента и остальной частью белковой молекулы. Именно это и является одной из причин высокой чувствительности и лабильности активного центра к различным воздействиям. 3. Ферменты, являясь истинными катализаторами, повышают скорость строго определенных химических реакций, которые в отсутствие их протекают очень медленно. При этом равновесие ускоряемых реакций не претерпевает изменений, в ходе реакций ферменты не расходуются и не изменяются, из чего следует, что для всех ферментов характерна специфичность их действия, т. е. каждый фермент атакует только вполне определенное вещество или действует на один и тот же тип связи в молекулах. Кроме того, специфичность проявляется в том, что ферменты действуют на один из возможных оптических изомеров. Например, тканевые ферменты человека и животных расщепляют только левовращающие аминокислоты. Специфичность действия ферментов имеет важное значение в 4.Одно из свойств ферментативного катализа -это обратимость, т. е. ферменты ускоряют как реакции синтеза, так и распада. Например, липаза катализирует синтез и распад жиров, пищеварительные ферменты при одних условиях расщепляют белки на свободные аминокислоты, а при других - синтезируют из их белок. Ряд ферментов существует в организме в неактивной форме Это- проферменты (зимогены), которые при определенных условиях переходят в активное состояние. Так, например, трипсиноген - неактивный профермент поджелудочной железы переходит в активный трипсин в тонком кишечнике под влиянием энте- рокиназы (активатора). Профермент пепсиноген активируется соляной кислотой желудка, липазой кишечника и желчными кислотами. Механизм перехода проферментов в активную форму различен. Так, например, тканевые протеиназы восстанавливаются веществами с SH-группой (цистеин, глютатион) или аскорбиновой кислотой. В других случаях это связано с разрушением связанного с ними ингибитора, отщеплением нескольких низкомолекулярных пептидов, переходом в восстановленное состояние и пр. Классификация ферментов. Современная классификация строится с учетом природы катализируемой ими реакции. Различают 6 классов ферментов. 1. Оксидоредуктазы - катализируют окислительно -восстановительные реакции (дегидрогеназы, лектроназы, цитохромы). 2. Трансферазы - катализируют перенос отдельных радикалов, частей молекул от одних соединений к другим (фосфотрансферазы, ацетилтрансферазы и т.д.). 3. Гидролазы - катализируют реакции расщепления сложных органических соединений в присутствии воды (амилазы, протеазы, эстеразы). 4. Лиазы - катализируют реакции отщепления от субстратов определенных групп с образованием двойных связей или присоединением отдельных групп по месту двойных связей (например, карбоксилаза). 5. Изомеразы - ферменты изомеризации, превращающие органические соединения в их изомеры (фосфогексизомеразы и пр.) 6. Лигазы - катализируют синтез сложных органических соединений из простых (например, карбоксилаза.) Механизм действия ферментов состоит в снижении энергии, активации, необходимой молекуле чтобы вступить в реакцию. Иммобилизация фермента - это методический прием, при котором молекулу биокатализатора включают в какую-либо фазу, отделенную от фазы свободного раствора, но способную обмениваться с ней молекулами субстрата, эффектора или ингибитора. Иммобилизованные ферменты имеют существенные преимущества по сравнению с нативными ферментными препаратами. Так, например, они легко отделимы от реакционной среды. Это дает возможность остановить реакцию в любой момент, получить продукт, незагрязненный катализатором, и использовать ферментный препарат многократно. Иммобилизованные ферменты технологичны, что определяется возможностью вести биотехнологический процесс непрерывно и регулировать скорость катализируемой реакции и выход продукта путем изменения скорости протока. Подбором соответствующих носителей и методов иммобилизации можно целенаправленно модифицировать такие свойства ферментов, как специфичность, рН- и температурозависимость, а также стабильность фермента при денатурирующих воздействиях. Успешное использование иммобилизованных ферментов в значительной мере определяется выбором подходящего сочетания носителя и метода иммобилизации, а также знанием кинетики реакций с участием таких катализаторов.
Носители для иммобилизации ферментов В качестве носителей для иммобилизованных ферментов могут использоваться различные вещества: природные и синтетические, органические и неорганические, высоко- и низкомолекулярные, но все они должны отвечать следующим требованиям: Органические высокомолекулярные носители по происхождению делят на 2 класса: Природные полимеры по химической природе подразделяются на белковые, полисахаридные и липидные. Наиболее широко в качестве носителей используются полисахариды, такие как целлюлоза, декстран, агароза и их производные. Их преимуществом являются доступность, наличие различных функциональных групп, высокая степень гидрофильности, недостатком - чувствительность к действию микроорганизмов и высокая стоимость. Целлюлоза- это поли-бета-глюкопираноза. Отличается высокой степенью гидрофильности и наличием большого количества ОН-групп. Последнее обстоятельство дает возможность получения модифицированных носителей введением различных заместителей. Известны ДЭАЭ-, КИ-, аминоэтил-, бромцианцеллюлоза и др. Механическую прочность целлюлоз повышают частичным гидролизом, разрушающим ее аморфные участки и позволяющим получать гранулированные формы. Недостаток этого носителя - чувствительность к действию кислот, щелочей и окислителей. Декстран- разветвленный полисахарид, содержащий остатки альфа-глюкозы. На его основе получены такие носители, как сефадексы (Швеция), молселект (Венгрия), образующие в водных растворах сильно набухающие гели, сжимающиеся при высушивании и обладающие высокой химической стойкостью. Легкость биодеградации позволяет применять их в медицине в качестве носителей лекарственных веществ. Агарозаявляется полимером альфа- и бета- галактозы. На ее основе получены различные марки таких полисахаридных носителей, как сефароза, биогель А и др. В последние годы в качестве носителей чаще используются природные полисахариды, полученные из морских водорослей - каррагинан, альгиновая кислота и ее соли. Каррагинан- гетерогенный полисахарид, содержащий, главным образом, эфиры галактозы и серной кислоты. В присутствии моновалентных катионов образует гели. Альгиновые кислоты и их соли- альгинаты состоят из связанных остатков маннуроновых кислот. В присутствии 2-х валентных катионов, особенно кальция, образуют гели. Преимущество этих носителей - получение геля при низких температурах, в мягких условиях, недостаток - невысокая механическая прочность. В работах последних лет сообщается об использовании в качестве носителей хитина и хитозана. Хитин- природный аминополисахарид, представляющий собой продукт переработки креветок и крабов, в состав наружного скелета которых он входит. По химической природе - это производное целлюлозы, в молекуле которой ОН-группа замещена ацетамидом. Хитин обладает пористой структурой, не растворяется в воде, разбавленных кислотах и щелочах, органических растворителях. Деацилирование его концентрированными растворами щелочей приводит к образованию хитозана, имеющего свободные аминогруппы и растворимого в минеральных кислотах и органических растворителях. Ферменты, иммобилизованные на этом носителе, высокоактивны, термостабильны, устойчивы к бактериальному заражению. Синтетические полимерные носители для иммобилизации ферментов создаются на основе: Известно, что in vivo большинство ферментативных реакций протекает на поверхности внутриклеточных или клеточных мембран, поэтому модель фермент-липид наиболее приближена к условиям функционирования энзима в клетке. Липидные носители используются в виде монослоев на различных поверхностях или в виде бислоев сферической формы (липосомы). Липосомы впервые были описаны А.Вэнгэмом в 1964 году, для их приготовления в основном используют фосфолипиды. Размер и форма липосом зависят от природы исходного субстрата, способа приготовления, кислотности среды и присутствия неорганических веществ. Простота получения и легкость регенерации иммобилизованного катализатора, возможность использования его in vivo, так как его свойства близки к таковым природных биомембран, способствовали широкому применению липосом в качестве носителей фермента и лекарственных препаратов. В настоящее время в качестве носителей внедряются синтетические аналоги липидов - поверхностно-активные вещества (ПАВ). Они имеют дифильную природу и содержат неполярную углеводородную часть и полярную головку, от содержания в которой различных функциональных групп различают 4 типа ПАВ: анионные, катионные, неионные и цвиттерионные. Многие из известных ПАВ являются продуктами крупнотоннажного производства, например фирмой "Serva" выпускается более 80 наименований только неионных ПАВ различного строения. Примером использования синтетических аналогов липидов в качестве носителей для иммобилизации являются синтетические моющие средства (СМС), такие как "Ока" и "Био", в состав которых входят смеси ПАВ с протеолитические ферментом. Неорганические носители - это матрицы на основе силикагеля, глины, керамики, природных минералов и их оксидов. Они могут быть пористыми и непористыми и применяются в виде шариков и монолита. Преимущества этих носителей - легкость регенерации и возможность придания им любой конфигурации.
Методы иммобилизации
Известны 2 группы методов иммобилизации ферментов: Физические методы Самым старым способом иммобилизации является физическая адсорбция. Впервые в 1916 году Нельсон и Грифин адсорбировали фермент - инвертазу на активированном угле и геле гидроксида алюминия. В основе этого способа - физическое или ионное взаимодействие фермента с носителем, который может быть неорганическим (кремнезем, пористое стекло, природные алюмосикаты, оксиды алюминия или титана и др.) или органическим веществом (полисахариды, коллаген, ионообменные смолы и др.). Проведение адсорбции очень простой процесс и заключается в контакте водного раствора фермента с носителем и отмывке не адсорбированного фермента. Более удобным способом является колоночный, когда раствор фермента прокачивают через колонку с носителем и последующее отмывание проводят в этой же колонке. В 1969 году в лаборатории Чибата (Япония) впервые была разработана технология и получена первая партия иммобилизованного фермента - аминооксидазы, расщепляющей ацетил - аминокислоты. Преимущества адсорбционной иммобилизации - простота методики, доступность и дешевизна сорбентов, возможность придания им любой формы. Недостаток - десорбция и ее последствия. Это связано с тем, что сорбция обратима, она зависит от рН и ионной силы раствора, температуры, удельной поверхности и пористости носителя, концентрации субстрата и т.д. Для уменьшения степени десорбции иммобилизацию проводят: Недостатков адсорбции удается избежать при иммобилизации ферментов в поры геля. Это тоже физический метод, заключающийся в том, что энзим как бы погружают в матрицу геля, отделяющего его от субстрата полупроницаемой оболочкой. Методически такая процедура выполняется различными способами: Гели, используемые для иммобилизации ферментов, содержат 50-90% воды, заполняющей пространство между полимерными цепями. Фермент, заключенный в матрицу геля, не может выйти наружу, во-первых, потому что расстояние между соседними цепями полимера меньше размеров его молекулы и, во-вторых, из-за наличия ионных и водородных связей фермента с носителем. Для иммобилизации ферментов применяются: В настоящее время широкое распространение получили сшитые гели на основе производных акриловой кислоты - полиакриламидные гели (ПААГ), которые были впервые использованы в качестве носителей в 1969 году Инманом и Динцисом. Включение фермента проводится на стадии синтеза полимера из мономера - акриламида в присутствии сшивающего агента - метилен-бис-акриламида. Образующийся блок геля с включенными в его матрицу молекулами фермента может быть подвергнут механическому воздействию для получения частичек разной величины и формы (шарики, волокна, пленки и т.д.). Преимущества ферментов, включенных в матрицу геля: Недостатки: Другие взаимодействия возникают между носителем, ферментом и субстратом при иммобилизации ферментов с использованием полупроницаемых оболочек: Химические методы При использовании химических методов иммобилизации между ферментами и носителем возникают ковалентные связи. В качестве носителей выступает неограниченный набор природных или синтетических материалов различного молекулярного веса. Ковалентные связи образуются за счет того, что ферменты, являясь веществами белковой природы, содержат на поверхности молекулы различные функциональные группы: гидроксильные, карбоксильные, тиоловые, амино-, имидазольные, гуанидиновые и т.д. Самые реакционные - это SH-группы, принимающие участие в реакциях окисления, ацилирования, алкилирования и др. Эти группы играют большую роль в стабилизации белка-фермента при помощи дисульфидных мостиков. Так как сульфгидрильные группы входят в активный центр многих ферментов, их не используют для иммобилизации, а, напротив, защищают. В качестве группы - мишени при химической иммобилизации чаще всего используют высокореакционные аминогруппы белка, которых достаточно много и роль которых в поддержании структуры фермента второстепенна. В некоторых случаях процесс иммобилизации протекает с участием третьего участника - сшивающего агента (би- или полифункциональный реагент) с образованием следующих конструкций: Пришивка методически осуществляется таким же образом как адсорбция, т.е. фермент и носитель смешиваются, в результате чего фермент адсорбируется на поверхности носителя. Но, в отличие от простой адсорбции, при химической иммобилизации между функциональными группами на поверхностях фермента и носителя образуются ковалентные связи, за счет которых фермент пришивается к носителю. Вшивка или ретикуляция ферментов в различные типы сеток осуществляется при помощи сшивающих агентов, от природы и количества которых зависит растворимость образующихся сетчатых структур, узлами в которых служат молекулы ферментов. Фактически, в этом случае фермент сам является носителем. Если тесное взаимодействие фермента с носителем нежелательно, т.к. ограничивает стерические и диффузионные процессы, то его можно предотвратить при помощи сшивающих молекул различной длины (“сшивка”). Таким образом, ковалентная иммобилизация ферментов позволяет получать препараты с заданными и контролируемыми свойствами. Но, вследствие дороговизны и сложности их получения, необходимости в обязательной защите активного центра энзима и т.д., этот метод используется , в основном, в лабораторной практике. В крупномасштабных производствах предпочтительна физическая иммобилизация. Выбор носителя и метода иммобилизации для каждого конкретного случая носит эмпирический характер и может контролироваться только экспериментально.
|