| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
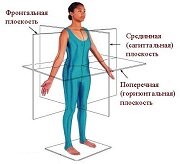
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Таким образом, ацетоацетат в норме выполняет роль источ-ника энергии для сердечной мышцы, скелетных мышц, мозга.
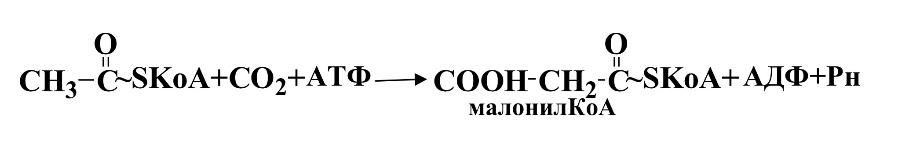
Голоданиеидиабет,ведущие к усиленному освобождению жир-ных кислот из тканевых депо и к снижению метаболизма углеводов в печени, приводят к образованию такого избытка кетоновых тел, что внепеченочные ткани не справляются с их утилизацией. Это приводит к накоплению кетоновых тел в крови(кетонемия), которые обладают свойствами кислот, что снижает рН и развивается метаболический аци-доз. При большом избытке кетоновых тел они выводятся почками,т.е. возникает кетонурия. В крайне тяжелых случаях ацетон выводится через легкие и может быть обнаружен в выдыхаемом воздухе БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ: СИНТЕЗ И РОЛЬ МАЛОНИЛ-КОА, ХАРАКТЕРИ-СТИКА ПАЛЬМИТАТСИНТЕТАЗНОГО КОМПЛЕКСА, РЕГУЛЯЦИЯ. ПУТИ ОБРАЗО-ВАНИЯ ЖИРНЫХ КИСЛОТ С БОЛЕЕ ДЛИННОЙ УГЛЕРОДНОЙ ЦЕПЬЮ И НЕНА-СЫЩЕННЫХ ЖИРНЫХ КИСЛОТ. Биосинтез жирных кислот можно рассматривать как процесс, складывающийся из 3 этапов.
I. Транспорт ацетил-КоА в цитозоль из митохондрий II. Образование малонилКоА
III. Конденсация этих молекул и их восстановление с образовани-ем высших насыщенных жирных кислот, главным образом пальмитино-вой.
I этап.Образование ацетил-КоА происходит в митохондриях,а ихмембрана непроницаема для ацетил-КоА.Перенос ацетильных групп происходит при помощи цитрата (цитратный челночный механизм).
ЩУК может вернуться в митохондрии с помощью своей трансло-казы, но чаще она восстанавливается до малата с участием малатдегид-рогеназы (МДГ).
Образующийся НАДФН2 используется в дальнейшем для синтеза жирных кислот.
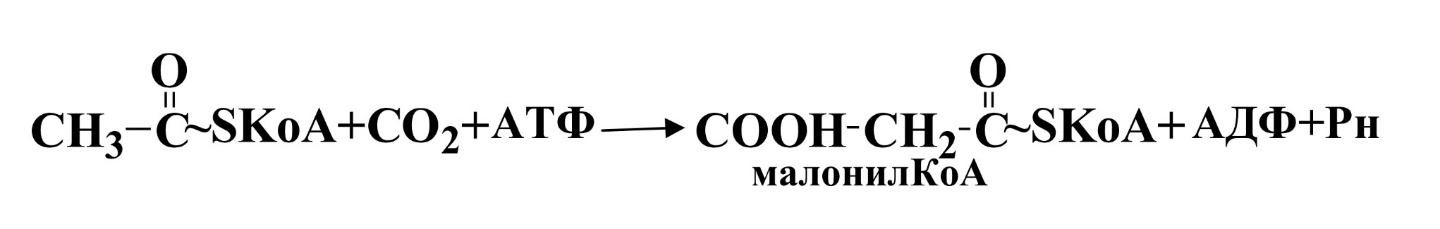
II этап.Ацетил-КоА карбоксилируется под действием ацетил-КоА-карбоксилазы, сложного фермента, коферментом которого служит витамин биотин.
Эта реакция лимитирует скорость всего процесса синтеза жирных кислот.
III этаппротекает при участиимультиферментного пальмитат- синтазного комплекса.Он состоит из двух полипептидных цепей.Ка-ждая полипептидная цепь содержит все 6 ферментов синтеза (трансаци-лаза, кетоацилсинтаза, кетоацилредуктаза, гидратаза, еноилредуктаза, тиоэстераза). Ферменты связаны между собой ковалентными связями, ацилпереносящий белок (АПБ) является также частью полипептидной цепи, но его функция связана только с переносом ацильных радикалов. В процессе синтеза важную роль играюттиогруппы. Одна из них при-надлежит 4-фосфопантотеину, входящему в состав АПБ (центральная) и вторая – цистеину кетоацилсинтазы (периферическая). Функциональ-ная единица синтеза состоит из половины одного мономера, взаимодей-ствующего с комплементарной половиной второго мономера,где цен-
тральная SH-группа одного мономера очень близка к периферической SH-группе другого. Т.е. на синтазном комплексе синтезируются одно-временно 2 жирные кислоты и только димер активен. Перенос субстрата от фермента к ферменту происходит при участии АПБ Реакции 3 этапа
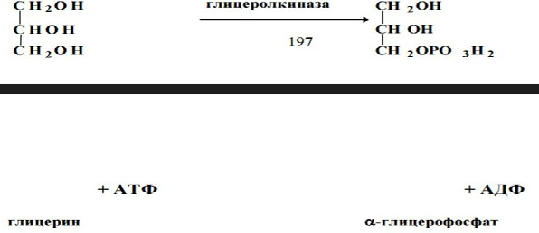
13. БИОСИНТЕЗ ТРИАЦИЛГЛИЦЕРИНОВ, ЛИЦЕРОФОСФОЛИПИДОВ. Триацилглицеринысинтезируются во многих органах и тканях, но наиболее важную роль в их синтезе играют печень, стенка кишечника, лактирующая молочная железа и жировая ткань. Для синтеза необходима активная форма глицерина – a-глицерофосфат и активная форма жирной кислоты – ацил-КоА. Активация глицерина может происходить двумя способами:1. В стенке кишечника и почках, печени есть активная глицерол- киназа:
2. В жировой ткани и мышцах активность этого фермента очень низкая и образование a-глицерофосфата связано с гликолизом:
a-Глицерофосфат, образованный любым из этих путей, взаимо- действует с двумя молекулами активированных жирных кислот (ацил- КоА) с образованием фосфатидной кислоты:
6. Синтез глицерофосфолипидов Биосинтез глицерофосфолипидов наиболее интенсивно происходит в печени, стенке кишечника, семенниках, молочной железе. Реакции синтеза локализованы в эндоплазматической сети. Синтез до образования фосфатидной кислоты происходит также, как и синтез триацилглицеринов.
14. ОБМЕН И ФУНКЦИИ ХОЛЕСТЕРИНА. ПРЯМОЙ И ОБРАТНЫЙ ТРАНСПОРТ ХО-ЛЕСТЕРИНА. БИОСИНТЕЗ ХОЛЕСТЕРИНА: ХИМИЗМ, РЕГУЛЯЦИЯ. Обмен холестерина. Транспорт холестерина 1. Обмен холестерина Функции холестерина: А – быстрообменивающийся (около 30 г холестерина); В – медленнообменивающийся (около 50 г холестерина); С – очень медленнообменивающийся (около 60 г холестерина). К первому пулу А следует отнести холестерин печени и других паренхиматозных органов, а также холестерин кишечной стенки и плазмы крови. Обновление холестерина этого пула происходит в среднем за 30 сут (1 г/сут). К третьему пулу (пул С) можно отнести холестерин головного и спинного мозга, нервов и холестерин соединительной ткани. Скорость обновления холестерина в белом веществе мозга исчисляется годами. Холестерин остальных органов и тканей составляет промежуточный медленнообменивающийся пул В. 2. Транспорт холестерина Прямой транспорт холестерина Рецепторы захвата ЛПНП
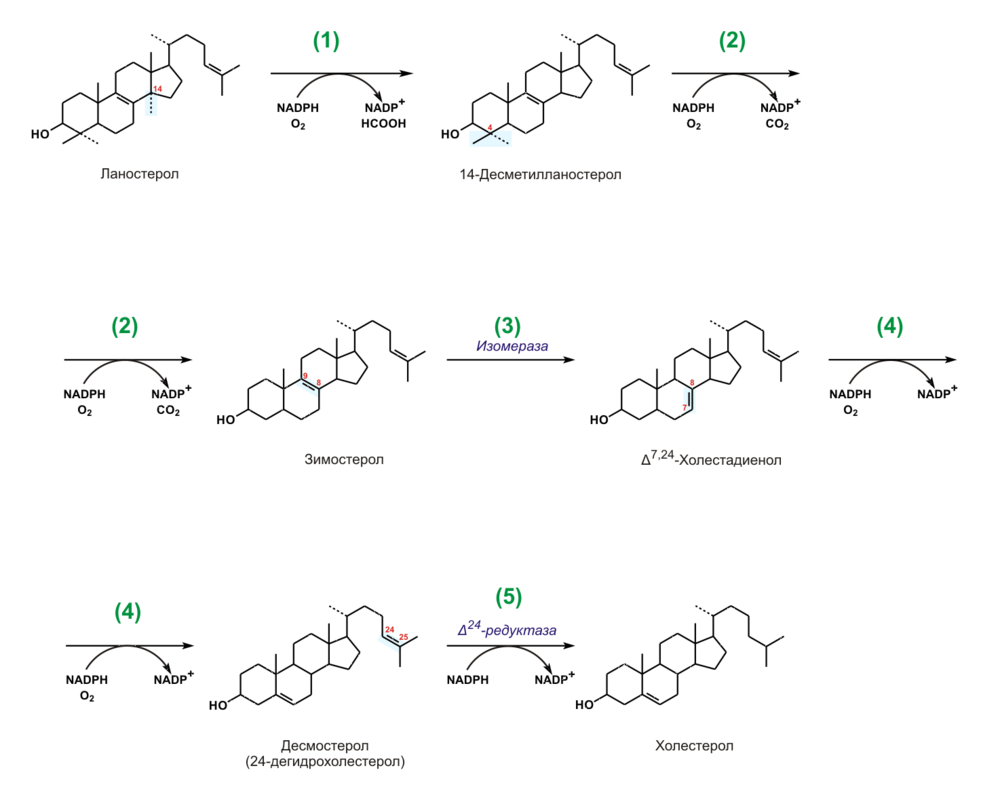
Поступивший холестерин клетка использует для построения мембран. Такой рецепторно-опосредованный путь поступления холестерина в клетку предохраняет ее от перегрузки холестерином, т.к.: Синтез холестерина Ланостерин превращается в мембранах гладкого эндоплазматического ретикулума в холестерин. Метильная группа при С14 окисляется, и образуется 14-десметилланостерин. 2. Затем удаляются ещё два метила при С4, и образуется зимостерол. 3. Далее двойная связь C8=С9 перемещается в положение С8=С7 и образуется Δ7,24-холестадиенол. 4. Двойная связь далее перемещается в положение С5=С6,образуется десмостерол. 5. После чего в боковой цепи восстанавливается двойная связь, и образуется холестерин. (Восстановление двойной связи в боковой цепи может, однако, происходить и на предшествующих стадиях биосинтеза холестерола.)
|