 ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение
Как определить диапазон голоса - ваш вокал
Игровые автоматы с быстрым выводом
Как цель узнает о ваших желаниях прежде, чем вы начнете действовать. Как компании прогнозируют привычки и манипулируют ими
Целительная привычка
Как самому избавиться от обидчивости
Противоречивые взгляды на качества, присущие мужчинам
Тренинг уверенности в себе
Вкуснейший "Салат из свеклы с чесноком"
Натюрморт и его изобразительные возможности
Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д.
Как научиться брать на себя ответственность
Зачем нужны границы в отношениях с детьми?
Световозвращающие элементы на детской одежде
Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия
Как слышать голос Бога
Классификация ожирения по ИМТ (ВОЗ)
Глава 3. Завет мужчины с женщиной
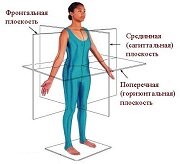
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
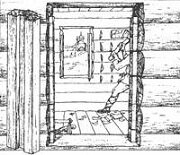 Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу. Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу.
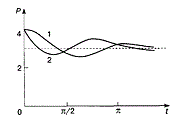 Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар. Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.
| Концепция Леонтьева - Фабри
Относительно становления и развития психики и поведения у животных существует целый ряд гипотез. Одну из них, касающуюся стадий и уровней развития психического отражения, от простейших животных и до человека, в своей книге "Проблемы развития психики" выдвигает А.Н. Леонтьев. Позднее, с учетом многих исследований, касающихся поведения, эта гипотеза была доработана и уточнена К.Э. Фабри. Поэтому рассматриваемую в нашем учебном курсе гипотезу развития психики принято называть концепцией Леонтьева - Фабри.
К.Э. Фабри считает, что как в пределах элементарной сенсорной, так и в пределах перцептивной психики следует выделить существенно различающиеся уровни психического развития: низший и высший, допуская при этом существование и промежуточных уровней. Важно отметить, что крупные систематические группы животных не всегда и не вполне укладываются в эти рамки. Это неизбежно, так как в пределах крупных таксонов, например классов, подтипов или типов, всегда имеются роды или виды, стоящие на смежных уровнях психического развития. Это можно объяснить тем, что качества высшего психического уровня всегда зарождаются на предшествующем уровне.
По мнению Фабри, расхождения между психологической и зоологической классификациями обусловлены тем, что морфологические признаки, на которых построена систематика животных, отнюдь не всегда определяют особенности и степень развития психической деятельности последних. Поведение животных представляет собой совокупность функций эффекторных органов животных. А в процессе эволюции именно функция первично определяет форму, строение организма, его систем и органов. Их строение и двигательные возможности лишь вторично определяют характер поведения животного и ограничивают сферу его внешней активности.
Этот диалектический процесс, однако, осложняется еще возможностями многопланового решения задач и компенсаторными процессами в области поведения. Это означает, что если животное в данных условиях лишено возможности решить биологически важную задачу одним путем, оно, как правило, имеет в своем распоряжении еще другие, резервные возможности. Так, одни эффекторы могут замениться другими, т.е. разные морфологические структуры могут служить для выполнения биологически однозначных действий. С другой стороны, одни и те же органы могут выполнять разные функции, т.е. осуществляется принцип мультифункциональности. Особенно пластичны морфофункциональные отношения в координационных системах, и, прежде всего, в центральной нервной системе высших животных.
Итак, с одной стороны, образ жизни определяет развитие приспособлений в эффекторной сфере, а с другой стороны, функционирование эффекторных систем, т.е. поведение, обеспечивает удовлетворение жизненных потребностей, обмена веществ в ходе взаимодействия организма с внешней средой.
При этом врожденное и приобретаемое поведение не являются последовательными ступенями на эволюционной лестнице, а развиваются и усложняются совместно, как два компонента одного единого процесса. Прогрессивному развитию инстинктивного, генетически фиксированного поведения соответствует прогресс в области индивидуально-изменчивого поведения. Инстинктивное поведение достигает наибольшей сложности как раз у высших животных, и этот прогресс влечет за собой развитие и усложнение у них форм обучения
######################## Психическая эволюция - предмет генетической психологии. Она занимается решением двух глобальных проблем - психической макроэволюцией и психической микроэволюцией. В первом случае речь идет о психической эволюции животных и людей в целом, а во втором - о психической эволюции того или иного вида животного или человека в отдельности. В первом случае мы имеем дело с филогенетической психологией, а во втором - с онтогенетической. А.Н.Леонтьев делил психическую эволюцию на четыре стадии - элементарной сенсорной психики, перцептивной, интеллектуальной и человеческой. 1. Стадия элементарной сенсорной психики. Под элементарной сенсорной психикой А.Н.Леонтьев имел в виду способность живых организмов к чувствительности или, что одно и то же, к ощущению. Ощущение, с его точки зрения, - это отправной пункт психической эволюции. «Итак, - писал он, - мы будем считать элементарной формой психики ощущение, отражающее внешнюю объективную действительность, и будем рассматривать вопрос о возникновении психики в этой конкретной его форме как вопрос о возникновении «способности ощущений», или, что то же самое, собственно чувствительности» Способностью к ощущению, по мнению ученого, обладают животные, обладающие нервными клетками. Но не только! Он усматривал способность к ощущению (а следовательно, элементарную психику) не только у тех животных, у которых появились нервные клетки, либо разбросанные по всему телу, как у полипов, либо уже объединенные в сцепления, как у других кишечнополостных - кораллов и медуз, но и у тех, у которых нервные клетки еще отсутствуют, как у губок. Их чувствительность обеспечивается на уровне протоплазмы. Чувствительность (или способность к ощущению) возникла не на голом месте. Она возникла из раздражимости, которая является общим свойством всей живой материи. Под раздражимостью обычно понимают способность организма реагировать на температурные, световые и т.п. физические изменения в окружающей среде. Если вслед за А.Н.Леонтьевым мы примем чувствительность за начало психики, то ее предтечу - раздражимость - очевидно, следует расценивать как предпсихику. Но где граница между раздражимостью (предпсихикой) и чувствительностью (психикой)? А.Н.Леонтьев писал: «Невозможность объективно различать между собою процессы чувствительности и раздражимости привела физиологию последнего столетия вообще к игнорированию проблемы этого различения. Поэтому часто оба эти термина - чувствительность и раздражимость - употребляются как синонимы» Отказаться от разграничения раздражимости и чувствительности означает не что иное, как отказаться от решения вопроса о возникновения психики и, более того, от решения вопроса о том, что такое психика вообще. Вот почему А.Н.Леонтьев стал подходить к вопросу о разграничении раздражимости и чувствительности со стороны решения проблемы психики как таковой. «Психика, - писал он, - есть свойство живых, высокоорганизованных материальных тел, которое заключается в их способности отражать своими состояниями окружающую их, независимо от них существующую действительность… Психические явления - ощущения, представления, понятия - суть более или менее точные и глубокие отражения, образы, снимки действительности Сущность психики, таким образом, составляет способность организма отражать внутри себя существующую действительность с помощью ощущений представлений, понятий. Исходя из отражательной природы психики вообще, А.Н.Леонтьев пришел к выводу о том, что ощущение как первичная форма психического отражения есть результат эволюционного развития раздражимости. Чувствительность - новая ступень в эволюции живой природы по сравнению с раздражимостью. Если раздражимость еще не оставляет отражательных «снимков» в организме, то чувствительность их оставляет в виде ощущений. Под чувствительностью при этом имеется в виду способность организма различать элементарные свойства тех предметов, с которыми он взаимодействует в своей жизни. Эволюция психики, таким образом, началась от элементарной формы отражения действительности - ощущения, но продвигалась она к ее самой сложной форме, которую А.Н.Леонтьев назвал «образом мира». Как ощущение (начальный пункт психогенеза), так и образ мира (конечный пункт психогенеза, присущий только человеку) суть разные формы отражательной деятельности живого организма. Эта деятельность должна рассматриваться, по убеждению А.Н.Леонтьева, в контексте всей жизнедеятельности организма, в контексте деятельности, направленной на приспособление к окружающей среде. Деятельность, о которой идет речь, есть не что иное как взаимодействие живого организма с внешним миром. Понятие деятельности выступает у А.Н.Леотьева как основополагающее. Не случайно его психологическую концепцию часто называют деятельностной. «Итак, - писал ее автор, - основной вывод, который мы можем сделать, заключается в том, что для решения вопроса о возникновении психики мы должны начинать с анализа тех условий жизни и того процесса взаимодействия, который ее порождает. Но такими условиями могут быть только условия жизни, а таким процессом - только сам материальный жизненный процесс» (С.36). А чуть дальше он дает определение жизни: «Жизнь есть процесс особого взаимодействия особым образом организованных тел» Подобный, деятельностный, подход к вопросу о возникновении психики позволил А.Н.Леонтьеву увидеть в психогенезе продолжение биогенеза, т.е. связать жизнь в единое эволюционное целое с психикой. А.Н.Леонтьев писал: «Если рассматривать какой-нибудь процесс взаимодействия в неорганическом мире, то оказывается, что оба взаимодействующих тела стоят в принципиально одинаковом отношении к этому процессу. Иначе говоря, в неорганическом мире невозможно различить, какое тело является в данном процессе взаимодействия активным (т.е. действующим), а какое - страдательным (т.е. подвергающимся действию)» . Так обстоит дело в неживой природе. А каким является отношение между телами в живой природе? «Совершенно другое положение мы наблюдаем в случае взаимодействия органических тел, - читаем мы у А.Н.Леонтьева. - Совершенно очевидно, что в процессе взаимодействия живого белкового тела с другим каким-нибудь телом, представляющим для него питательное вещество, отношение обоих этих тел к самому процессу взаимодействия будет различным. Поглощаемое тело является предметом воздействия живого тела и уничтожается как таковое ). В живой природе, таким образом, субъекты действия взаимодействуют с их объектами. Вот почему переход природы в живую можно представить как переход от бессубъектно-безобъектной формы взаимодействия материальных тел к его субъектно-объективной форме. Вот как об этом писал А.Н.Леонтьев: «Иначе это можно выразить так: переход от тех форм взаимодействия, которые свойственны неорганическому миру, к формам взаимодействия, присущим живой материи, находит свое выражение в факте выделения субъекта, с одной стороны, и объекта - с другой» . Наличие субъекта действия и его объекта в живой природе указывает нам на наличие в ней деятельности: «Те специфические процессы, которые осуществляют то или иное жизненное, т.е. активное, отношение субъекта к действительности, мы будем называть в отличие от других процессов процессами деятельности» . Понятия деятельности и жизни у А.Н.Леонтьева, таким образом, есть эволюционно синхронные понятия. Раздражимость, с точки зрения ученого, была самой элементарной формой деятельности живого организма, выражающейся в способности организма отвечать специфическими процессами на то или другое жизненно значимое воздействие Ученый различал два вида раздражимости. Первый из них связан с реакцией организма, направленной на поддержание жизни, а второй - не направленный на поддержание жизни. Чувствительность - результат развития раздражимости первого вида. Переход «поддерживающей» раздражимости к чувствительности знаменовал собою возникновение психики у животных. Развитие чувствительности у животных в свою очередь закреплялось появлением у них и совершенствованием органов чувствительности, эволюционировавших постепенно в органы чувств. Последние появились в результате все большей специализации первых. В основе этого перехода лежала необходимость во все более адекватном отражении организмом той предметной действительности, с которой он вступал во все более и более сложные отношения. 2. Стадия перцептивной психики. Развитие органов чувств, сопровождаемое развитием нервной системы, привело животных ко второй стадии их психической эволюции - стадии перцептивной психики. В отличие от первой, она характеризуется способностью животного отражать предметную действительность не только в виде ощущений, но и в виде представлений. Последние позволили им схватывать не отдельные свойства окружающих вещей, что доступно ощущениям, а вещи в целом. А.Н.Леонтьев писал: «Деятельность животных на самой ранней, первой стадии развития психики характеризуется тем, что она отвечает тому или иному отдельному воздействующему свойству… Следующая… стадия… характеризуется способностью отражения внешней объективной действительности уже не в форме отдельных элементарных ощущений, вызываемых отдельными свойствами или их совокупностью, но в форме отражения вещей» . На второй, перцептивной, стадии психогенеза «окружающая действительность отражается животным в форме более или менее расчлененных образов отдельных вещей» . Очевидно, следует согласиться с К.Э.Фабри, который возразил А.Н.Леонтьеву в том, что перцептивной, целостной, психикой обладают только наземные позвоночные. Опираясь на более современные наблюдения, К.Э.Фабри расширил область перцептивной психики за счет рыб, а также высших беспозвоночных - моллюсков, ракообразных, высших насекомых и др А.Н.Леонтьев принижал перцептивные возможности животных за счет преувеличения роли элементарной сенсорной психики, которую он приписывал по существу всем животным, кроме высших позвоночных, начиная с наземных. Развитие перцептивной психики А.Н.Леонтьев в первую очередь связывал с совершенствованием дистантных органов чувств - зрения и слуха. Это совершенствование позволило животным подняться на новую ступень их психической эволюции, которая характеризуется освоением ими новой формы психического отражения - представлений. Представления и есть целостный образ предмета в сознании. 3. Стадия интеллекта. «Психика большинства млекопитающих животных, - писал А.Н.Леонтьев, - остается на стадии перцептивной психики, однако наиболее высокоорганизованные из них поднимаются еще на одну ступень развития» Эту новую ступень психического развития животных он назвал стадией интеллекта и стал приписывать ее по преимуществу человекообразным обезьянам. Очевидно, и здесь был прав К.Э.Фабри, возражая А.Н.Леонтьеву в том, что интеллектом наделены только человекообразные обезьяны. Им обладают и другие животные, если судить о наличии интеллекта у животных и людей по их способности оперировать понятиями, т.е. обобщенными представлениями - скажем, о дереве вообще, о собаке вообще и т.д. Интеллектуальные способности животных и до сих пор недооцениваются в зоопсихологии. А.Н.Леонтьев видел проявление интеллекта у обезьян в первую очередь в их способности к самостоятельному научению использовать предметы, оказавшиеся рядом, в качестве орудий. Так, обезьяны научаются пользоваться палкой, чтобы с ее помощью добраться до пищи. Они способны также использовать для этой цели ящики, складывая их друг на друга. И т.д. Отношение обезьян к подобным орудиям отличается от человеческого: «Когда в руках обезьяны палка выполнила свою функцию, она снова превращается для нее в безразличный предмет. Поэтому животные не хранят своих «орудий», не передают их из поколения в поколение. Они, следовательно, не способны выполнять той, по выражению Дж.Бернала, «аккумулирующей» функции, которая свойственная культуре» 4. Стадия человеческого сознания. «Итак, - писал А.Н.Леонтьев, - интеллектуальное поведение, которое свойственно высшим млекопитающим и которое достигает особенно высокого развития у человекообразных обезьян, представляет собой ту верхнюю границу развития психики, за которой начинается история развития уже совсем другого, нового типа, свойственная только человеку, - история развития человеческого сознания» Три предшествующие стадии психической эволюции - элементарной сенсорной психики, перцептивной и интеллектуальной - А.Н.Леонтьев квалифицировал как докультурные (а следовательно, как дочеловеческие). В качестве ведущего фактора психической эволюции человека он рассматривал фактор культуры. Вот почему его картину психической эволюции следует расценивать как культурологическую. Вот какой обобщенный образ культуры А.Н.Леонтьев давал в своем основном труде: «В своей деятельности люди не просто приспосабливаются к природе. Они изменяют ее в соответствии со своими развивающимися потребностями. Они создают предметы, способные удовлетворить их потребности, и средства для производства этих предметов - орудия, а затем и сложнейшие машины. Они строят жилища, производят одежду и другие материальные ценности. Вместе с успехами в производстве материальных благ развивается и духовная культура людей; обогащаются их знания об окружающем мире и о самих себе, развивается наука и искусство» В своей психической эволюции человек сумел так существенно оторваться от животных по той причине, что он стал создавать продукты материальной и духовной культуры. Стремление к их улучшению выступало в истории людей в качестве стимула для развития их интеллекта, поскольку любой продукт культуры первоначально моделируется в сознании его творца. Развитие культуры, таким образом, есть главный фактор развития человеческого мышления. Основным показателем интеллектуального развития у человека является его способность к творчеству, благодаря которой и создаются новые культурные ценности. Однако прежде, чем кто-либо овладеет этой способностью, он должен освоить уже имеющиеся культурные достижения. Процесс их освоения есть не что иное как процесс очеловечивания. А.Н.Леонтьев писал: «…каждый отдельный человек учится быть человеком. Чтобы жить в обществе, ему недостаточно того, что ему дает природа при его рождении. Он должен еще овладеть тем, что было достигнуто в процессе исторического развития человеческого общества. Перед человеком целый океан богатств, веками накопленных бесчисленными поколениями людей - единственных существ, населяющих нашу планету, которые являются созидателями. Человеческие поколения умирают и сменяют друг друга, но созданное ими переходит к следующим поколениям, которые в своих трудах и борьбе умножают и совершенствуют переданные им богатства - несут дальше эстафету человечества» Еще более выразительно об очеловечивании он писал таким образом: «Человек не рождается наделенным историческими достижениями человечества. Достижения развития человеческих поколений воплощены не в нем, не в его природных задатках, а в окружающем его мире - в великих творениях человеческой культуры. Только в результате процесса присвоения человеком этих достижений, осуществляется в ходе его жизни, он приобретает подлинно человеческие свойства и способности; процесс этот как бы ставит его на плечи предшествующих поколений и высоко возносит над всем животным миром Усвоить достижения предшествующей культуры и внести в нее свой вклад - вот в чем видел А.Н.Леонтьев смысл человеческой жизни. Вот почему он беспокоился о создании общественных условий, позволяющих успешно реализовывать этот смысл в жизни каждого человека. Великий психолог не сомневался в эволюционных возможностях человека, но он видел проблему в создании условий, позволяющих реализовывать эти возможности. Он писал: «Итак, действительная проблема заключается не в способности или неспособности людей овладеть достижениями человеческой культуры, сделать их достижениями своей личности и внести в них свой вклад. Действительная проблема заключается в том, чтобы каждый человек и все люди, все народы получили практическую возможность вступить на путь ничем не ограниченного развития. Это и есть та великая цель, которая стоит сейчас перед прогрессивным человечеством. Цель эта достижима. Но она достижима в условиях, которые способны реально освободить людей от бремени материальной нужды, уничтожить уродующее их разделение умственного и физического труда, создать систему воспитания, обеспечивающую всестороннее и гармоническое их развитие, дающее возможность каждому творчески участвовать во всех проявлениях человеческой жизни» (С.435). Сердце обливается кровью, когда читаешь эти золотые слова! Уж очень далеко отодвинули от нас великую цель, о которой говорил А.Н.Леонтьев. 32.Уровни развития психики животных. Эволюция психики составляет часть общего процесса эволюции животного мира и совершалась по закономерностям этого процесса. Повышение общего уровня жизнедеятельности организмов, усложнение их взаимоотношений с окружающим миром приводило в ходе эволюции к необходимости все более интенсивного контактирования со все большим числом предметных компонентов среды, к совершенствованию передвижения среди этих компонентов и ко все более активному обращению с ними. Только такое прогрессивное изменение поведения могло обеспечить возрастающее потребление необходимых для жизни компонентов среды, равно как все более успешное избегание неблагоприятных или вредных воздействий и опасностей. Однако для всего этого требовалось существенное совершенствование ориентации во времени и пространстве, что и достигалось прогрессом психического отражения. У животных уровень развития психики связан с уровнем их отношений с окружающей средой. Выделяют следующие типы взаимосвязей животного с средой: 1. Взаимосвязи с территорией (главная сфера): - территория проживания и ее состав; - по каким признакам животное ориентируется; - функциональные зоны (где питается, охотится, пережидает опасность. Некоторые виды способны подстраивать местность под себя, некоторые виды могут замечать новое в местности. 2. Взаимосвязи в сфере питания: - сведения о рационе животного. 3. Взаимосвязи в репродуктивной сфере: - как образуется пара; - как животное устраивает место рождения потомства; - забота о детенышах и взаимосвязи с ними. 4. Взаимосвязи в сфере общения с сородичами: - образ жизни (одиночный, семейный, стадный); - структура группы, иерархия; - организация отношений между животными (функциональное деление в стае, не иерархия); - язык животных. 5. Взаимосвязи в сфере общения с человеком: - способность животного устанавливать контакт с человеком, отвечает ли оно на попытки человека контактировать и каким образом; - способность животного освоить чуждый ему язык, научиться тому, что выходит за рамки его видотипичного поведения (эта способность является показателем уровня развития психики животного). Схема определения уровня развития психики: - Животное. - Его вид, структура, особенности строения, нервная система. - Его взаимодействие со средой и другими животными. - Пластичность поведения в естественной среде, способность приспосабливаться к изменениям. - Как животное проявляет себя в экспериментах. У животных уровень развития психики связан с уровнем их отношения с окружающей средой. Выделяют: 1)Элементарную сенсорную психику(низший и высший уровни); 2)Перцептивную психику(низший и высший уровень. Согласно представлениям Леонтьева, стадия элементарной сенсорной психики характеризуется примитивными элементами чувствительности, не выходящими за пределы простейших ощущений. Низший уровень стадии элементарной сенсорной психики, на котором находятся простейшие и низшие многоклеточные организмы, живущие в водной среде, характеризуется тем, что здесь в достаточно развитом виде представлена раздражимость - способность живых организмов реагировать на биологически значимые воздействия среды повышением уровня своей активности, изменением направления и скорости движений. Чувствительность как способность реагировать на биологически нейтральные свойства среды и готовность к научению методом условных рефлексов еще отсутствует. Двигательная активность животных еще не имеет поискового, целенаправленного характера.
На высшем уровне развития этой стадии психики у животных отмечается выделение специализированного органа, осуществляющего сложные манипулятивные движения организма с предметами внешнего мира. Таким органом у низших животных являются челюсти. Они заменяют им руки, которые есть только у человека и некоторых высших живых существ. Челюсти сохраняют свою роль как основной орган манипуляций и исследования окружающего мира в течение длительного периода эволюции, вплоть до освобождения для этой цели передних конечностей животного. 33.Низший уровень элементарной сенсорной психики. Анатомофизиологические особенности животных. Характеристика психики. Низший уровень сенсорной психики. На низшем уровне психического развития находится довольно большая группа животных; среди них встречаются и такие животные, которые стоят еще на грани животного и растительного мира (жгутиковые), а с другой стороны, и сравнительно сложно устроенные одноклеточные и многоклеточные животные. 1. Простейшие. К наиболее типичным представителям рассматриваемой здесь группы животных относятся простейшие. Организм представителей этого типа состоит из единственной клетки, обеспечивающей все жизненные потребности животного. Филогенез простейших шел фактически параллельно развитию многоклеточных животных, что нашло свое отражение в формировании у простейших аналогов систем органов, так называемых органелл.
На низшей ступени развития жизни у простейших одноклеточных животных наблюдается разнообразное поведение. Под микроскопом в капле воды можно видеть, как движутся, питаются, размножаются и погибают амёбы и инфузории. Сложность движений этих организмов поразительна. О трудностях изучения жизнедеятельности простейших животных проф. В. А. Вагнер остроумно и справедливо пишет: "В термине "простейшие" больше иронии, чем правды. Изучение их жизни не проще, чем изучение сложных организмов".
Движения простейших отличаются большим разнообразием, причем у представителей этого типа встречаются способы локомоции, присущие только им и совершенно отсутствующие у многоклеточных животных. Это, например, своеобразный способ передвижения амеб при помощи "переливания" плазмы из одного участка тела в другой. Другие представители простейших, грегарины, передвигаются своеобразным "реактивным" способом - путем выделения из заднего конца тела слизи, толкающей животное вперед. Существуют и простейшие, пассивно парящие в воде.
Однако большинство простейших передвигаются активно с помощью особых структур, производящих ритмичные движения, - жгутиков или ресничек. Они представляют собой плазматические выросты, совершающие колебательные, вращательные или волнообразные движения. Жгутиками, длинными волосовидными выростами обладают примитивные простейшие - жгутиковые, получившие свое название благодаря этому образованию. С помощью жгутиков тело животного (например, эвглены) приводится в спиралевидное поступательное движение. Более сложным двигательным аппаратом являются реснички, покрывающие в большом числе тело инфузорий. Как правило, реснитчатый покров располагается неравномерно, реснички достигают на разных участках тела различной длины, образуют кольцевидные уплотнения (мембранеллы) и т.п. Так, например, инфузории-стилонихии при помощи этих своеобразных органелл способны не только плавать, но и "бегать" по твердому субстрату, причем как вперед, так и назад. Жгутики и реснички приводятся в движение сокращениями миофибрилл, которые образуют волоконца, мионемы, соответствующие мышцам многоклеточных животных. У большинства простейших они являются основным двигательным аппаратом, причем имеются они даже у наиболее примитивных представителей типа жгутиковых. Мионемы располагаются в строгом порядке, чаще всего в виде колец, продольных нитей или лент, а у высших представителей и в виде специализированных систем. Сложные системы мионем позволяют простейшим производить не только простые сократительные движения тела, но и достаточно разнообразные специализированные локомоторные и не локомоторные движения.
У тех простейших, у которых нет мионем (у амеб, корненожек, споровиков и некоторых других простейших), сократительные движения совершаются непосредственно в цитоплазме. Таким образом, еще задолго до появления мышц перемещение животного в пространстве совершается путем сокращений. Именно сократительная функция, которую у простейших осуществляют мионемы, а у многоклеточных мышцы, обеспечивала все разнообразие и всю сложность двигательной активности животных на всех этапах филогенеза.
Элементарные движения простейших иначе называют кинезами. Типичным примером кинеза является ортокинез - поступательное движение с переменной скоростью. Если, например, на определенном участке существует температурный градиент (перепад температур), то движения инфузории-туфельки будут тем более быстрыми, чем дальше животное будет находиться от места с оптимальной температурой. Следовательно, здесь интенсивность поведенческого (локомоторного) акта непосредственно определяется пространственной структурой внешнего раздражителя.
В отличие от ортокинеза при клинокинезе имеет место изменение направления передвижения. Это изменение не является целеустремленным, а носит характер проб и ошибок, в результате которых животное в конце концов попадает в зону с наиболее благоприятными параметрами раздражителей. Частота и интенсивность этих изменений зависят от интенсивности воздействующего на животное отрицательного раздражителя (или раздражителей). С ослаблением силы действия этого раздражителя уменьшается и интенсивность клинокинеза. В данном случае животное также реагирует на градиент раздражителя, но не увеличением или уменьшением скорости передвижения, как при ортокинезе, а поворотами оси тела, т.е. изменением вектора двигательной активности.
Таким образом, осуществление наиболее примитивных инстинктивных движений - кинезов - определяется непосредственным воздействием градиентов интенсивности биологически значимых внешних факторов. Роль внутренних процессов, происходящих в цитоплазме, заключается в том, что они дают поведенческому акту "первый толчок", как и у многоклеточных животных. Ориентация. Уже на примерах кинезов мы видели, что градиенты внешних раздражителей выступают у простейших одновременно как пусковые и направляющие стимулы. Особенно наглядно это проявляется при клинокинезах. Однако изменения положения животного в пространстве еще не являются здесь подлинно ориентирующими, поскольку они носят ненаправленный характер. Для достижения полного биологического эффекта клинокинетические, как и ортокинетические, движения нуждаются в дополнительной коррекции, позволяющей животному более адекватно ориентироваться в окружающей его среде по источникам раздражения, а не только менять характер движения при неблагоприятных условиях.
Ориентирующими элементами являются у представителей рассматриваемого типа и у других низших беспозвоночных, стоящих на данном уровне психического развития, простейшие таксисы. В ортокинезах ориентирующий компонент - ортотаксис - проявляется в изменении скорости передвижения без изменения его направления в градиенте внешнего раздражителя. В клинокинезах этот компонент называется клинотаксисом и проявляется в изменении направления движения на определенный угол. Под таксисами понимают генетически фиксированные механизмы пространственной ориентации двигательной активности животных в сторону благоприятных (положительные таксисы) или в сторону от неблагоприятных (отрицательные таксисы) условий среды. Так, например, отрицательные термотаксисы выражаются у простейших, как правило, в том, что они уплывают из зон с относительно высокой температурой воды, реже - из зон с низкой температурой. В результате животное оказывается в определенной зоне термического оптимума (зоне предпочитаемой температуры). В случае ортокинеза в температурном градиенте отрицательный ортотермотаксис обеспечивает прямолинейное удаление от неблагоприятных термических условий. Если же имеет место клинокинетическая реакция, то клинотаксис обеспечивает четкое изменение направления передвижения, ориентируя тем самым случайные клинокинетические движения в градиенте раздражителя (в нашем примере - в термическом градиенте).
Зачастую клинотаксисы проявляются в ритмичных маятникообразных движениях (на месте или при передвижении) или в спиралевидной траектории плывущего животного. И здесь имеет место регулярный поворот оси тела животного (у многоклеточных животных это может быть и только часть тела, например голова) на определенный угол.
Клинотаксисы обнаруживаются и при встрече с твердыми преградами. Например, наткнувшись на твердую преграду (или попав в зону с другими неблагоприятными параметрами среды), инфузория-туфелька останавливается, у нее изменяется характер биения ресничек, и она отплывает немного назад. После этого инфузория поворачивается на определенный угол и снова плывет вперед. Это продолжается до тех пор, пока она не проплывет мимо преграды (или не минует неблагоприятную зону).
В приведенных примерах описывались реакции простейших (в данном случае инфузорий) на температуру и тактильный раздражитель (прикосновение). Речь шла, следовательно, о термо- и тигмотаксисах, в последнем случае - об отрицательном тигмоклинотаксисе, возникающем в ответ на сильное тактильное раздражение (соприкосновение с твердой поверхностью объекта).
Если же, наоборот, туфелька натыкается не на твердое препятствие, а на мягкий объект (например, растительные остатки, фильтровальная бумага), она реагирует иначе: при такой слабой тактильной стимуляции инфузория останавливается и прикладывается к этой поверхности так, чтобы максимальный участок тела соприкасался с поверхностью объекта (положительный тигмотаксис). Аналогичная картина наблюдается и при воздействиях других модальностей на направление движения, т.е. положительный или отрицательный характер реакции зависит от интенсивности раздражения. Как правило, простейшие реагируют на слабые раздражения положительно, на сильные - отрицательно, но в целом простейшим больше свойственно избегать неблагоприятных воздействий, нежели активно искать положительные раздражители.
Возвращаясь к тигмотаксисам, важно отметить, что у инфузорий обнаружены специальные рецепторы тактильной чувствительности - осязательные "волоски", которые особенно выделяются на переднем и заднем концах тела. Эти образования служат не для поиска пищи, а только для тактильного обследования поверхностей объектов, с которыми животное сталкивается. Раздражение этих органелл и приводит в описанном примере к прекращению кинетической реакции.
Особенностью тигмотаксисной реакции является то, что она часто ослабевает, а затем и прекращается после прикасания к объекту максимальной поверхностью тела: приставшая к объекту туфелька в возрастающей мере начинает реагировать на иной раздражитель и все больше отделяется от объекта. Затем, наоборот, вновь возрастает роль тактильного раздражителя и т.д. В результате животное совершает возле объекта ритмичные колебательные движения.
Четко выражена у туфельки и ориентация в вертикальной плоскости, что находит свое выражение в тенденции плыть вверх (отрицательный геотаксис - ориентация по силе земного притяжения). Поскольку у парамеции не были обнаружены специальные органеллы гравитационной чувствительности, было высказано предположение, что содержимое пищеварительных вакуолей действует у нее наподобие органов равновесия - статоцистов высших животных. Обоснованность такого толкования подтверждается тем, что туфелька, проглотившая в опыте металлический порошок, плывет уже не вверх, а вниз, если над ней поместить магнит. В таком случае содержимое вакуоли (металлический порошок) уже давит не на нижнюю ее часть, а, наоборот, на верхнюю, чем, очевидно, и обусловливается переориентация направления движения животного на 180°.
Кроме упомянутых, таксисные реакции установлены у простейших также в ответ на химические раздражения (хемотаксисы), электрический ток (гальванотаксисы) и др. На свет часть простейших реагируют слабо, у других же эта реакция выражена весьма четко. Так, фототаксисы проявляются у некоторых видов амеб и инфузорий. Светочувтсвтиельность. В отличие от инфузорий, у многих жгутиковых, особенно у эвглены, положительный фототаксис выражен весьма четко. Биологическое значение этого таксиса не вызывает сомнений, так как аутотрофное питание эвглены требует солнечной энергии. Эвглена плывет к источнику света по спирали, одновременно, как уже упоминалось, вращаясь вокруг собственной оси. Это имеет существенное значение, так как у эвглены, как и у некоторых других простейших, сильно и положительно реагирующих на свет, имеются хорошо развитые аналоги фоторецепторов. Это пигментные пятна, иногда снабженные даже отражающими образованиями, позволяющими животному локализовать световые лучи. Продвигаясь к источнику света описанным образом, эвглена поворачивает к нему то "слепую" (спинную) сторону, то "зрячую" (брюшную). И каждый раз, когда последняя (с незаслоненным участком "глазка") оказывается обращенной к источнику света, производится корректировка траектории движения путем поворота на определенный угол в сторону этого источника. Следовательно, движение эвглены к свету определяется положительным фотоклинотаксисом, причем в случае попадания ее под воздействие двух источников света попеременное раздражение фоторецептора то слева, то справа придает движению эвглены внешнее сходство с тропотаксисным поведением двустороннесимметричных животных, обладающих парными глазами.
"Глазки" описаны и у других жгутиковых. Особую сложность фоторецепция достигает у одного из представителей динофлагеллят, у которого имеются уже аналоги существенных частей глаза многоклеточных животных, пигментное пятно снабжено не только светонепроницаемым экраном (аналог пигментной оболочки), но и светопроницаемым образованием в форме сферической линзы (аналог хрусталика). Такой "глазок" позволяет не только локализовать световые лучи, но и собирать, в известной степени фокусировать их. Пластичность поведения простейших. Итак, поведение простейших и в моторной и в сенсорной сфере у ряда видов уже достигает известной сложности.
У организмов, лишенных нервной системы, обнаружен целый ряд форм адаптивного поведения, напоминающих обучение.
Сенсибиллизация. Сенсибилизацией называется повышение чувствительности организма к воздействию какого-либо агента, способствующей модификации поведения. Ярким примером такого рода может служить так называемое обучениеинфузорий.
В обычных условиях инфузории-туфельки передвигаются в воде как бы толчками. Их движения носят хаотичный характер. Никаких закономерностей и ни малейшей целенаправленности в поведении инфузорий не наблюдается.
Если же инфузорию перенести в небольшую емкость, имеющую форму круга, глубиной не больше 1 мм и диаметром 3-5 мм, то ее поведение резко изменится. Сначала она будет хаотично двигаться по сосуду, изредка натыкаясь на его стенки. Однако уже через 3-4 мин поведение инфузории изменится: путь ее станет прямолинейным, и вскоре она начнет описывать правильную геометрическую фигуру, форма которой зависит от формы сосуда. Так, в круглом аквариуме это будет почти правильной восьмиугольник; в квадратном - квадрат, расположенный косо по отношению к стенкам аквариума; в пятиугольном сосуде - пятиугольник; в шестиугольном - шестиугольник и т.д. При этом, будучи перенесенными в сосуд другой формы, инфузории в течение некоторого времени продолжают двигаться по предыдущей траектории. Подобных опытов было проведено множество. Почти всегда инфузории демонстрировали высокую способность к обучению. Выработанные у них реакции по своему характеру и по способу их образования напоминали условные рефлексы высших животных. Некоторые исследователи их так и называли: "условные рефлексы простейших". Более тщательно проведенные исследования полностью опровергли представления о высоких способностях инфузорий. Грубая ошибка произошла из-за незнания особенностей врожденных форм поведения туфелек. Наблюдения за инфузориями показали, что хаотические движения сохраняются у них только до тех пор, пока они находятся в культуральной жидкости, где всегда много углекислого газа и мало кислорода. Когда ту же жидкость наливают в экспериментальный сосуд тонким слоем, она обогащается кислородом. В таких условиях движения инфузорий становятся прямолинейными, а при столкновении с препятствием туфелька отскакивает от него под углом 20°. Поэтому после помещения инфузории в широкий и мелкий сосуд путь инфузории начинает повторять его конфигурацию. Подобная реакция на изменения внешней среды представляет собой типичную сенсибилизацию первого типа, но никак не обучение.
Привыкание простейших. Примером подобного элементарного накопления индивидуального опыта служит привыкание. Напомним, что под привыканием понимается прекращение реакции на постоянно действующий раздражитель. По принятой системе классификации обучения его относят к типу неассоциативного обучения.
Способность к выработке привыкания обнаружена у самых примитивных организмов. Из одноклеточных существ для подобных исследований чаще всего используют исключительно крупную разноресничную инфузорию спиростомум амбигуум. Эта инфузория достигает в длину 2 мм и хорошо видна невооруженным глазом.
Если к поверхности крохотного аквариума, где находится спиростомум, прикоснуться кончиком карандаша, вызвав колебание пленки поверхностного натяжения, а вслед за ней и толщи воды, все находящиеся там инфузории мгновенно, как по команде, прекратят движение и съежатся в комочек. Испуг от неожиданного вторжения в их маленький мир скоро пройдет, тела инфузорий вытянутся, и они как ни в чем не бывало продолжат свое движение. Притрагиваясь раз за разом к поверхности аквариума, удается приучить его обитателей меньше бояться безобидного воздействия. Скоро инфузории перестанут полностью сжиматься и будут быстрее возобновлять обычное движение. Проявив настойчивость, можно приучить спиростомумов совершенно не обращать внимание на сотрясение воды, не сжиматься в комочек и не прекращать движения.
Безусловно, в лаборатории инфузорий "дрессируют", не прибегая к помощи карандаша. Их приучают к легкой вибрации аквариума, создаваемой специальным прибором. Если включать вибрацию с интервалами в 7 сек., то уже через 1-10 мин станет заметно, что инфузории не так сильно боятся ее. Продолжая тренировку, можно через 13-47 мин добиться полного привыкания.
У простейших привыкание весьма недолговечно и не поддается тренировке. Если через час после выработки у инфузорий привыкания проверить его сохранность, то окажется, что спиростомумы полностью отвыкли от вибрации и снова реагируют на нее обычной оборонительной реакцией. Чтобы они привыкли опять, потребуется столько же предъявлений раздражителя, сколько было сделано первый раз. Сравнение шести последовательных сеансов тренировки привыкания, проведенных с часовым интервалом, не обнаруживает какого-либо ускорения его восстановления к концу опыта. У инфузории стентнор память значительно лучше. Эта крупная сидячая инфузория, напоминающая крохотную воронку, способна 3-6 ч помнить о том, что слабого механического раздражения бояться не нужно.
Кроме вибрации, у спиростомумов удалось выработать привыкание к прикосновению и электрическому воздействию. И в этих случаях "привычка" не бояться внезапного действия раздражителя сохранялась 30-50 мин, и при попытке ее восстановить облегчающего влияния предыдущей тренировки не было заметно (цит. по Сергеев, 1986). Говоря о привыкании простейших, важно подчеркнуть, что этот феномен проявляется не только у ресничных инфузорий, но и у более простых одноклеточных, например амебы. Таким образом, как показали многочисленные эксперименты, приобретенное поведение простейших обладает свойствами, общими с таковыми у животных, имеющих нервную систему, и одновременно несет черты более примитивной организации.
Л.Г. Воронин (1968) относит привыкание простейших к несигнальной форме индивидуального приспособления.
Сам по себе факт наличия у животных, лишенных даже зачатков нервной системы, элементов поведения, напоминающих процесс обучения, представляет исключительный интерес для общих представлений об эволюции психики. 2. Кишечнополостные. У представителей типа кишечнополостных уже отмечаются зачатки нервной системы.
В своей простейшей форме она встречается у гидр и актиний, представляя собою нервную сеть, состоящую из разбросанных нервных клеток с отростками, переплетающимися между собой. Такая нервная сеть не имеет особых центров, и возбуждение проходит по всем направлениям. Подобная первичная нервная система называется рассеянной, или диффузной.
У некоторых кишечнополостных, в связи с усложнением строения организма, нервная ткань начинает концентрироваться в определенных местах тела. У медуз, например, на краю зонтика, где находятся щупальцы и органы чувств, нервная ткань образует кольцевой тяж. Отсюда во все стороны отходит сеть нервных клеток с длинными отростками.
Наблюдения и опыты показывают, что кишечнополостные животные довольно тонко различают механические, химические, световые и температурные раздражители. В опытах Леба актинии втягивали щупальцами кусочки мяса и переваривали их, в то же время они отталкивали бумажные трубки, по величине и форме сходные с мясом.
Рассеянная нервная система не всегда приводит к единству действия всех частей организма как целого. При слабых раздражителях, например, наблюдаются движения отдельных щупальцев актиний. Точно так же реагируют отделенные от организма части гидры, сохранившие нервные клетки. Длительно действующее раздражение постепенно распространяется по всему организму. Вопрос о способности кишечнополостных формировать условные рефлексы в настоящее время остается открытым. Результаты немногочисленных экспериментальных попыток выработки условных реакций у различных видов этого типа не выявили свойств приобретенных реакций, которые можно было бы определить как условнорефлекторные.
В то же время неассоциативное обучение по типу привыкания у кишечнополостных осуществляется лучше и сохраняется дольше, чем у простейших.
Например, стебельчатая гидра, как и инфузории, пугается вибрации. Однако память у нее надежнее: через час после выработки еще удается обнаружить привыкание, но через сутки никаких следов от него не остается. Голодная гидра хватает любой объект, коснувшийся ее щупалец, и даже может заглотить несъедобную добычу. Поймав первый раз крохотную кварцевую песчинку, гидра под ее тяжестью валится на бок. В таком положении животное находится довольно долго. В лупу видно, с каким трудом она вытаскивает щупальца из-под придавившей их песчинки. Когда ей наконец удается освободиться от добычи и принять нормальную позу, можно кинуть новую песчинку. Гидра непременно соблазнится и схватит очередное подношение. Животное долго будет "охотиться" на несъедобный кварц, но время освобождения от него станет постепенно сокращаться, а 25-35-ю песчинку животное уже не станет удерживать. Это не усталость. Наткнувшуюся на нее дафнию гидра непременно поймает и отправит по назначению. Привыкание к песчинке сохраняется от 40 мин до нескольких часов. Даже через сутки можно еще обнаружить следы привыкания: второй раз научить гидру не трогать несъедобную добычу оказывается легче.
В результате выяснилось, что при интервалах, сравнимых со временем сохранения следа после однократного привыкания, наблюдается ярко выраженное ускорение привыкания от опыта к опыту. Этот результат представляет интерес, поскольку именно по степени выраженности тренированности у животных различного уровня филогенеза (или нейронных систем различной сложности) при различных интервалах между приложениями можно судить о степени консолидации следа и, возможно, определить четкий критерий различия памяти кратковременной и долговременной - основных функциональных механизмов поведения.
Таким образом, прогресс в развитии функциональных механизмов поведения у кишечнополостных по сравнению с простейшими заключается в появлении нового свойства привыкания - тренированности (см. 3. Плоские черви. Низшие ресничные черви, или турбеллярии, имеют значительно более совершенную нервную систему по сравнению с ранее описанными группами животных.
Одна из замечательных филогенетических особенностей ресничных червей, к которым относятся планарии, заключается в том, что они представляют уровень "перехода" диффузной нервной сети в концентрированную систему. У ресничных червей впервые в эволюции нервные элементы концентрируются на переднем конце тела, т.е. появляются зачатки цефализации. Интегрирующее значение нервной системы на этой стадии филогенеза выражается в регулировании церебральным ганглием важных функций организма.
Церебральный ганглий планарий состоит в основном из мелких, малодифференцированных клеток. Основной особенностью гистологического строения ганглия служит наличие в нем большого количества мышечных и паренхиматозных клеток, выполняющих, по-видимому, нейросекреторные функции. Только отдельные ("гигантские") клетки могут быть с большей степенью уверенности классифицированы как нервные.
Таким образом, появившаяся впервые в эволюции у планарий, цефализация представлена здесь в своей примитивной форме и к ней приурочена приобретаемая в индивидуальной жизни организма примитивная форма нестойких условных рефлексов. Вопрос о выработке истинных классических условных рефлексов у низших (ресничных) червей, представителями которых являются планарии, долгое время считался дискуссионным.
В результате статистической обработки результатов исследования авторы пришли к выводу о возможности выработки классических условных рефлексов у планарий. Временная связь у всех планарий была нестойкой, количество положительных реакций из каждых 10 сочетаний условного сигнала с безусловным редко превышало 50%.
Из анализа экспериментального материала следует, что условные рефлексы у изученных пресноводных планарий недостаточно стойки, не обладают всеми качествами классических условных рефлексов. Они характеризуются следующими признаками, общими для экологически различных видов животных: непрочностью в течение одного опыта, непрочностью от опыта к опыту (упрочение не наступало даже после 335 сочетаний условного сигнала с безусловным), угасанием реакций после 200-300 сочетаний, несмотря на подкрепление. Перечисленные свойства условных рефлексов не являются отражением индивидуальных особенностей отдельных видов, так как они характерны для животных с различной экологией. Таким образом, подобные реакции можно отнести к категории лишь примитивных нестойких условных рефлексов, свойственных животным определенного уровня филогенетического развития.
У планарий, предпочитающих держаться в затемненных местах, удалось выработать привыкание к свету, но животное, прошедшее начальный "курс" обучения, не кажется поумневшим. И все же плоские черви способнее, чем одноклеточные и кишечнополостные животные. Предъявляя им слабые раздражители, которых они и так не пугаются, у них можно образовать привыкание к более сильным. 5. Общая характеристика низшего уровня элементарной сенсорной психики (по Фабри, 1976). Итак, на низшем уровне элементарной сенсорной психики поведение животных выступает в достаточно разнообразных формах, но все же с примитивными проявлениями психической активности. Простейшим свойственна элементарная форма психического отражения - ощущение, т.е. чувствительность в собственном смысле слова. Как утверждает Леонтьев, даже низший уровень психического отражения не является низшим уровнем отражения вообще, существующего в живой природе, в частности растениям присуще допсихическое отражение, при котором имеют место лишь процессы раздражимости.
Степень и качества психического отражения определяются тем, насколько развиты способности к движению, пространственно-временной ориентации и к изменению врожденного поведения. У простейших встречаются разнообразные формы передвижения в водной среде только на самом примитивном уровне инстинктивного поведения - кинезов. Ориентация поведения осуществляется только на основе ощущений и ограничена элементарными формами таксисов, позволяющими животному избегать неблагоприятные внешние условия.
Это значит, что поисковая фаза инстинктивного поведения у них еще крайне плохо развита и лишена сложной, многоэтапной структуры. Во многих случаях эта фаза вообще отсутствует. Во всем этом проявляется не только исключительная примитивность инстинктивного поведения на данном уровне, но и предельная скудность содержания психического отражения. Как уже отмечалось, в отдельных случаях у простейших встречаются и положительные элементы пространственной ориентации. Так, например, амеба в состоянии находить пищевой объект на расстоянии до 20-30 микрон. Зачатки активного поиска жертвы существуют, очевидно, и у хищных инфузорий. Однако во всех этих случаях положительные таксисные реакции еще не носят характера подлинного поискового поведения, поэтому эти исключения не меняют общую оценку поведения простейших, а тем более характеристику низшего уровня элементарной сенсорной психики в целом. На этом уровне дистантно распознаются преимущественно отрицательные компоненты среды; биологически "нейтральные" же признаки положительных компонентов, как правило, еще не воспринимаются на расстоянии как сигнальные. Таким образом, психическое отражение на низшем уровне своего развития выполняет в основном сторожевую функцию и отличается поэтому характерной "однобокостью". Что касается пластичности поведения простейших, то и здесь простейшие обладают лишь самыми элементарными возможностями. Это вполне закономерно: элементарному инстинктивному поведению может соответствовать лишь элементарное неассоциативное научение, представленное наиболее примитивными формами.
Тем не менее при всей своей примитивности поведение простейших является все же достаточно сложным и гибким, в тех пределах, которые необходимы для жизни в своеобразных условиях микромира. Эти условия отличаются рядом специфических особенностей, и этот мир нельзя себе представить как просто во много раз уменьшенный макромир. В частности, среда микромира является менее стабильной, чем среда макромира, что проявляется, например, в периодическом высыхании маленьких водоемов, С другой стороны, непродолжительность жизни микроорганизмов и частая смена их поколений делают излишним - развитие более сложных форм накопления индивидуального опыта. Как уже отмечалось, простейшие не являются однородной группой животных, и различия между их разными формами очень велики. Высшие представители этого типа во многих отношениях развивались в своеобразных формах неклеточного строения параллельно низшим многоклеточным беспозвоночным животным. В результате высокоразвитые простейшие проявляют подчас даже более сложное поведение, чем некоторые многоклеточные беспозвоночные, также стоящие на низшем уровне элементарной сенсорной психики. К этому же уровню относятся и некоторые представители типа хордовых, например асцидии. Это служит наглядным подтверждением отмеченной выше закономерности: психологическая классификация не вполне совпадает с зоологической, так как одни представители одной и той же таксономической категории могут еще находиться на более низком психическом уровне, а другие - уже на более высоком. Билет 34. Высший уровень элементарной сенсорной психики. Анатомофизиологические особенности животных. Характеристика психики. Высший уровень элементарной сенсорной психики (по Фабри, 1976). Следующий, высший уровень стадии элементарной сенсорной психики, которого достигают живые существа типа иглокожих, кольчатых червей и брюхоногих моллюсков, характеризуется появлением первых элементарных ощущений, а также органов манипулирования в виде щупальцев и челюстей. Наиболее изученными из них являются кольчатые черви, к которым относятся живущие в морях многощетинковые черви (полихеты), малощетинковые черви ( олигохеты), наиболее известным представителем которых является дождевой червь, и пиявки. Характерным признаком их строения является внешняя и внутренняя метамерия: тело состоит из нескольких, большей частью идентичных, сегментов, каждый из которых содержит "комплект" внутренних органов, в частности пару симметрично расположенных ганглиев с нервными комиссурами, в результате нервная система кольчатых червей имеет вид "нервной лестницы".
На этом уровне развития психики находятся и низшие хордовые, которые вместе с позвоночными составляют тип хордовых. К низшим хордовым относятся оболочники и бесчерепные. Оболочники, или асцидии - морские животные, часть которых ведет неподвижную жизнь. Бесчерепные представлены всего двумя семействами с тремя родами мелких морских животных, наиболее известное из которых - ланцетник.
Изменчивость поведения животных, находящихся на этом уровне развития психики дополняется появлением способности к приобретению и закреплению жизненного опыта. На этом уровне уже существует чувствительность. Двигательная активность совершенствуется и приобретает характер целенаправленного поиска биологически полезных и избегания биологически вредных воздействий.
Виды приспособительного поведения, приобретаемые в результате мутаций и передаваемые из поколения в поколение благодаря естественному отбору, оформляются в качестве инстинктов.
Возникновение нервной системы беспозвоночных. Нервная система впервые появляется у низших многоклеточных беспозвоночных. Возникновение нервной системы - важнейшая веха в эволюции животного мира, и в этом отношении даже примитивные многоклеточные беспозвоночные качественно отличаются от простейших. Наличие нервной ткани способствует резкому ускорению проводимости возбуждения: в протоплазме скорость проведения возбуждения не превышает 1-2 микрон в секунду, но даже в наиболее примитивной нервной системе, состоящей из нервных клеток, она составляет 0,5 метра в секунду!
Нервная система существует у низших многоклеточных в весьма разнообразных формах: сетчатой (например, у гидры), кольцевой (медузы), радиальной (морские звезды) и билатеральной. Билатеральная форма представлена у низших плоских червей и примитивных моллюсков еще только сетью нервных клеток, располагающейся вблизи поверхности тела, в которой выделяются более мощным развитием несколько продольных тяжей. По мере своего прогрессивного развития нервная система погружается под мышечную, ткань, продольные тяжи становятся более выраженными, особенно на брюшной стороне тела. Одновременно все большее значение приобретает передний конец тела, появляется голова а вместе с ней и головной мозг - скопление и уплотнение нервных элементов в переднем конце. Наконец, у высших червей центральная нервная система уже вполне приобретает типичное строение "нервной лестницы", при котором головной мозг располагается над пищеварительным трактом и соединен двумя симметричными коммисурами ("окологлоточное кольцо") с расположенными на брюшной стороне подглоточными ганглиями и далее с парными брюшными нервными стволами. Существенными элементами являются здесь ганглии, поэтому подобную нервную систему называют ганглионарной, или "ганглионарной лестницей". У некоторых представителей данной группы например, пиявок, нервные стволы сближаются настолько, что получается "нервная цепочка".
От ганглиев отходят мощные проводящие волокна, которые и составляют нервные стволы. В гигантских волокнах нервные импульсы проводятся значительно быстрее благодаря их большому диаметру и малому числу синаптических связей (мест соприкосновения аксонов одних нервных клеток с дендритами и клеточными телами других клеток). Головные ганглии достигают лучшего развития у более подвижных животных, обладающих и наиболее развитыми рецепторными системами.
Зарождение и эволюция нервной системы обусловлены необходимостью координации разнокачественных функциональных единиц многоклеточного организма, согласования процессов, происходящих в разных частях его при взаимодействии с внешней средой, обеспечения деятельности сложно устроенного организма как единой целостной системы. Только координирующий и организующий центр, каким является центральная нервная система, может обеспечить гибкость и изменчивость реакции организма в условиях многоклеточной организации.
цефализации, т.е. обособление головного конца организма и сопряженное с ним появление головного мозга. Только при наличии головного мозга возможно подлинно централизованное "кодирование" поступающих с периферии сигналов и формирование целостных "программ" врожденного поведения, не говоря уже о высокой степени координации всей внешней активности животною.
Разумеется, уровень психического развития зависит не только от строения нервной системы. Так, например, близкие к кольчатым червям коловратки также обладают, как и те, билатеральной нервной системой и мозгом, а также специализированными сенсорными и моторными нервами. Однако, мало отличаясь от инфузории размером, внешним видом и образом жизни, коловратки очень напоминают последних также поведением и не обнаруживают более высоких психических способностей, чем инфузории. Этот пример показывает, что ведущим для развития психической деятельности является не общее строение, а конкретные условия жизнедеятельности животного, характер его взаимоотношений и взаимодействий с окружающей средой. 1. Кольчатые черви. Среди большой группы кольчатых червей, являющихся эволюционными потомками плоских червей, особое место занимают представители класса олигохета - дождевые черви, на которых проводились основные опыты, связанные с изучением их реакций на разнообразные агенты среды и с выработкой условных рефлексов. У червей нервные узлы (ганглии) расположены вдоль всего тела в виде симметричной цепочки. Каждый узел состоит из грушевидных клеток и густого сплетения нервных волокон. От клеток отходят нервные волокна к мышцам и к внутренним органам (двигательные волокна). Под кожным покровом червя расположены чувствительные клетки, которые соединяются своими отростками (чувствительные волокна) с нервными узлами. Нервная система подобного типа называется цепочечной, или ганглиозной. Тело дождевого червя состоит из ряда сегментов-члеников. Каждый сегмент имеет свой собственный нервный узел и может отвечать на раздражение, будучи совершенно отделен от всего остального тела. Но все узлы соединены между собой перемычками, и организм действует как целое. Головной узел нервной системы расположен в верхней части головы, получает и перерабатывает наибольшее количество раздражений. Он устроен значительно сложнее, чем все остальные узлы нервной системы червя.
Движения кольчатых червей. Итак, двигательная активность кольчатых червей отличается большим многообразием и достаточной сложностью. Обеспечивается это сильно развитой мускулатурой, состоящей из двух слоев: внешнего (подкожного), состоящего из кольцевых волокон, и внутреннего, состоящего из мощных продольных мышц. Последние простираются, несмотря на сегментацию, от переднего до заднего конца туловища. Ритмичные сокращения продольной и кольцевой мускулатуры кожно-мышечного мешка обеспечивают движения: червь ползет, вытягивая и сокращая, расширяя и сужая отдельные части своего тела. Так, у дождевого червя вытягивается и сужается передняя часть тела, затем то же самое происходит последовательно со следующими сегментами. В результате по телу червя пробегают "волны" сокращений и расслаблений мускулатуры.
У кольчатых червей впервые в эволюции животного мира появляются подлинные парные конечности. У них на каждом сегменте имеется по паре выростов, служащих органами передвижения, получивших название параподий, которые снабжены специальными мышцами, двигающими их вперед или назад. Зачастую параподии имеют ветвистое строение. Каждая ветвь снабжена опорной щетинкой и, кроме того, венчиком из щетинок, имеющих у разных видов различную форму. От параподин отходят и щупальцевидные органы тактильной и химической чувствительности. Особенно длинными и многочисленными последние являются на головном конце, где на спинной стороне располагаются глаза (одна или две пары), а в ротовой полости или на особом (выпячиваемом) хоботке-челюсти. В захвате пищевых объектов могут участвовать и нитевидные щупальца на головном конце червя.
У ряда многощетинковых и всех малощетинковых червей параподии редуцированы (отсюда и название последних), остались лишь посегментно расположенные пучки щетинок. Так, у дождевого червя на каждом сегменте находится по четыре пары очень коротких, неразличимых невооруженным глазом щетинок, которые, однако, наподобие параподий служат для передвижения животного: являясь достаточно крепкими подвижными рычагами, они обеспечивают вместе с сокращениями кожно-мышечного мешка поступательное движение червя. С другой стороны, растопыривая свои щетинки и упираясь ими в грунт, дождевой червь настолько прочно фиксирует свое тело в земле, что практически невозможно вытащить его оттуда в неповрежденном виде. У некоторых других малощетинковых червей щетинки развиты значительно сильнее и представлены в большем количестве.
Поведение кольчатых червей. Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная масса ведет придонный образ жизни в
|